Книги
Мотивированный мозг. Высшая нервная деятельность и естественно-научные основы общей психологии
В период пребывания крысы на педали когерентность падает, животное получает подкрепление и становится совершенно пассивным. Непосредственно перед уходом впервые возрастает когерентность негативного пункта и моторной области коры: возбуждение негативных структур готово к трансформации в двигательную реакцию избегания. После ухода животного с педали процент случаев достоверного возрастания когерентности возвращается к исходным цифрам. Только следы эмоционально-негативного состояния все еще обнаруживаются при сопоставлении электрической активности негативного пункта с двигательной областью коры.
Механизм обратной условной связи объясняет не только целенаправленность внешне реализуемых действий, но и природу таких явлений, как, например, феномен избирательного внимания. Наряду с условно-рефлекторным воспроизведением эффекта первого в сочетанной паре раздражителя тоническая обратная условная связь «может ограничиться одним лишь повышением возбудимости центральных структур последнего», настроив соответствующий анализатор к селекции строго определенных стимулов (Асратян, 1974, с. 14).
Концепция обратных условных связей позволяет объяснить классические и инструментальные (оперантные) условные рефлексы с единых теоретических позиций (Руис, 1984). «Благодаря работам ученика Павлова Асратяна, – пишут И. Гормезано и Р. Тейт, – гипотеза двусторонней условной связи стала центральным пунктом для павловского подхода к объяснению как классических, так и инструментальных условных рефлексов» (Gormezano, Tait, 1976). Обратным условным связям придается важное значение в трактовке целого ряда явлений, обнаруженных при изучении высшей нервной деятельности человека (Костандов, Важнова, Генкина, Захарова, Иващенко, Погребинский, 1984).
Итак, мы имеем веские основания рассматривать взаимодействие доминанты и условного рефлекса в качестве функциональной единицы индивидуально приобретаемого поведения. Уместно поставить вопрос: в какой мере эта теоретическая схема может быть «наложена» на морфофункциональную организацию реально существующего мозга?
Как было показано выше (см. гл. 3), экспериментальный и теоретический анализ генеза эмоциональных реакций человека и высших животных привел нас к выводу о решающем значении двух факторов, детерминирующих любое поведение. Мы имеем в виду наличие актуализированных потребностей и возможность их удовлетворения благодаря взаимодействию с внешней средой. Значимость стимулов, поступающих из внешней среды, зависит от их отношения к потребностям, имеющимся у организма, причем все эти стимулы можно разделить на две основные категории: на стимулы с высокой вероятностью их подкрепления факторами, непосредственно удовлетворяющими ту или иную потребность, и на стимулы с низкой вероятностью подкрепления. Среди актуальных потребностей, в свою очередь, выделяются наиболее острые доминирующие потребности, требующие первоочередного удовлетворения, и субдоминантные потребности, динамически сосуществующие с доминантной или конкурирующие с ней.
Экспериментальные данные, полученные в нашей лаборатории, а также почерпнутые из литературы, показали, что именно таким представлениям об организации поведения соответствует взаимодействие четырех мозговых образований, играющих решающую роль в оценке поступающих из внешней среды сигналов и выборе реакций. К ним относятся передние отделы новой коры, гиппокамп, миндалина и гипоталамус.
Поскольку в гипоталамусе были обнаружены структуры, связанные с актуализацией потребностей и с возникновением эмоций, можно сказать, что гипоталамус участвует и в самых ранних стадиях организации поведения и на более поздних его этапах, где окончательно оформляется внешне реализуемый ответ.
Ранняя (доминантная) стадия выработки нового условного рефлекса характеризуется широким вовлечением структур лимбической системы (Morrell, Barlow, Brazier, 1960), причем изменения нейронной активности и медленных потенциалов возникают в этих структурах раньше, чем в новой коре (Rebert, 1972; Linseman, Olds, 1973). Важнейшая роль в инициировании поведения принадлежит гипоталамусу (Судаков, 1971), поскольку двустороннее разрушение латерального гипоталамуса вызывает общее расстройство произвольного поведения (акинезию и каталепсию), которое включает афагию, но не ограничивается ею (Stricker, Andersen, 1980). Для таких животных характерны симптомы малоподвижности, вялости, застывания в одной и той же подчас необычной позе, т. е. признаки своеобразного «безволия». Хотя гипоталамус находится как бы у «истоков» поведенческого акта, его деятельность вторична по отношению к потребностно-мотивационным механизмам нижележащих отделов головного мозга и представляет сравнительно высокий уровень интеграции. Полагают, что в гипоталамусе нет собственно «вегетативных центров», а его влияние на вегетатику реализуется через эмоциогенные структуры, интегрирующие двигательные и вегетативные компоненты эмоций (Поляков, Талан, Черниговский, 1978).
В гипоталамусе обезьян обнаружены нейроны, реагирующие на вид только воды или только пищи, причем латентный период нейронов, реагирующих на вид пищи, равен 150–200 мс, а латентный период нейронов, реагирующих на прием пищи, составляет 300 мс (Rolls, Rolls, 1982). Стимуляция латерального гипоталамуса усиливает активность нейронов ядра одиночного пути, связанных со вкусовыми стимулами, и снижает активность нейронов, связанных с тепловой или механической чувствительностью языка (Matsuo, Shimizu, Kusano, 1984).
Степень голодового возбуждения оказывается решающим фактором для вовлечения структур гипоталамуса в процесс обучения и в реакции на внешние объекты, способные удовлетворить пищевую потребность. Только у голодных обезьян в латеральном гипоталамусе найдены нейроны, отвечавшие на вид и запах пищи, на вид шприца с глюкозой или скорлупу ореха (Mora, Rolls, Burton, 1976), причем введение глюкозы понижало реактивность нейронов на эти натуральные пищевые стимулы (Burton, Rolls, Burton, 1976). Чувствительные к введению глюкозы нейроны гипоталамуса вовлекаются в процесс пищедобывательного поведения: их активность резко возрастала у обезьян за 2–0,8 с перед нажатием на рычаг для получения пищи (Nishrino, Ono, Sasaki, Muramoto, 1979). Чувствительные к глюкозе нейроны связаны с норадренергической системой. Их импульсная активность подавлялась во время нажатий на рычаг для получения пищи. Нейроны, нечувствительные к введению глюкозы, напротив, усиливали свою активность при выполнении двигательных реакций. Это позволяет рассматривать нейроны первого типа как принадлежащие к побуждающей, мотивирующей поведение системе. Нейроны второго типа уместнее связать с выполнением двигательного акта или с предвосхищением предстоящего подкрепления (Aou, Oomura, Nishino, 1983). Полагают, что нейроны латерального гипоталамуса преимущественно участвуют в дискриминации пищевых и непищевых объектов и в инициировании пищедобывательного движения (Опо, 1981). Часть чувствительных к глюкозе нейронов изменяют свою импульсную активность в ответ на условный сигнал, большинство – на подкрепление пищей (Aou, Oomura, 1984).
Нейроны латерального гипоталамуса обезьян отвечают через 150–200 мс после открывания заслонки для подачи пищи, в то время как активность нейронов бледного шара изменяется только спустя 300 мс вместе с электромиографическими признаками двигательной реакции (Rolls, Roper-Hall, Sanghera, 1977). Аналогичная последовательность событий наблюдается и в процессе выработки нового условного рефлекса. Изменение частоты разрядов нейронов гипоталамуса возникает после 10–20 сочетаний звука с пищей, а поведенческие признаки нового рефлекса – только после 40–50 проб (Olds, 1973).
Исследования механизмов жажды и морфинизма у крыс показали, что нейроны латерального гипоталамуса активируются возникновением потребности (драйвом) и тормозятся ее удовлетворением (подкреплением). В медиальном гипоталамусе наблюдаются противоположные эффекты. Нейроны базальных отделов переднего мозга крыс весьма избирательно реагируют на значимые для животного объекты, будь то сексуальный партнер (медиальная преоптическая область), вода (латеральная перегородка), рука экспериментатора (латеральная преоптическая область) и т. п. (Mink, Sinnamon, Adams, 1983).
По сравнению с гипоталамусом функции второго мозгового образования, относимого к разряду «мотивационных», – ядра миндалевидного комплекса – в еще большей мере зависят от факторов внешней среды и наличной ситуации удовлетворения актуальных потребностей. Регуляция веса тела, потребление пищи и воды, ответ на уровень глюкозы в крови и продолжительность пищевой депривации существенно не изменяются после повреждения миндалины. Ее сохранность важнее для аппетита, для различения вкусной и невкусной, новой и знакомой пищи, т. е. для ответов на экстероцептивные пищевые стимулы, в том числе после короткой пищевой депривации, когда в организме еще не возникли метаболические сдвиги (Schoenfeld, Hamilton, 1981). Крысы с разрушением базолатерального ядра миндалины теряют способность избегать употребления неприятного для интактных крыс раствора (Rolls, Rolls, 1982). При выработке условнорефлекторного вкусового отвращения зарегистрирована реакция нейронов миндалины в ответ на условный сигнал. Возможно, что именно в миндалине происходит конвергенция вкусовых (условных) и висцеральных подкрепляющих сигналов (Буреш, Бурешова, 1979).
По-видимому, за счет миндалины животные, переставшие реагировать на тканевую жажду и падение уровня глюкозы в крови после разрушения латерального гипоталамуса, продолжают отвечать на условные сигналы воды и пищи. Базолатеральная часть миндалины связана с влиянием прошлого опыта утоления жажды, а не с «тканевой жаждой» (Rolls, Rolls, 1973). Разрушение вентральной части медиального ядра нарушает пищевые и особенно питьевые инструментальные условные рефлексы, не сказываясь сколько-нибудь существенно на потреблении воды и пищи (Korczynski, Fonberg, 1976). Аналогичные последствия повреждения миндалины наблюдаются и в сфере оборонительных реакций, где амигдалэктомия нарушает поведение, вызванное страхом, и не влияет на порог болевого раздражения (Ursin, 1965; Reeves, Martin, Ghiselli, 1977). Преимущественная связь функций миндалины с внешними, а не с внутренними мотивирующими стимулами хорошо согласуется с особенностями ее морфологических связей. Миндалина имеет мощные пути к гипоталамусу и более слабые – от него наряду с развитыми путями от лобных и височных зон неокортекса. Эта система (неокортекс – миндалина – гипоталамус) функционально формируется в процессе онтогенеза и играет важную роль в индивидуальных особенностях поведения (Чепурнов, Чепурнова, 1981).
Старая кортикомедиальная часть миндалины получает афферентные волокна от гипоталамуса, таламуса, продолговатого мозга, перерабатывающих висцеральные и ноцицептивные сигналы. Базолатеральная часть связана со структурами конечного мозга и таламуса. Таким образом, существуют два афферентных входа в миндалину: со стороны височной коры и со стороны ствола через задние ядра таламуса. Прямой путь более древний. Путь через кору эволюционирует параллельно с развитием неокортекса (Гончар, 1984). Исследование пространственной организации биопотенциалов новой коры, гиппокампа и миндалины у собак показало, что при упроченном инструментальном пищевом условном рефлексе возрастают когерентные функции дельта– и тета-диапазонов электрической активности в парах: миндалина – фронтальная кора, миндалина – сенсомоторная кора. Миндалина и гиппокамп взаимодействуют слабо, что, по-видимому, отражает их различную роль в организации поведения (Алексанов, Вайнштейн, Преображенская, 1984).
Суммируя имеющиеся данные о функциях миндалины и ее роли в организации поведения, П. Глур приходит к выводу о том, что «главный дефект, вызываемый повреждением амигдалы, можно описать как расстройство мотивационного механизма, который в норме позволяет выбрать поведение, приобретенное в данной ситуации» (Gloor, 1960, р. 1416). Миндалина оказывается необходима для перестройки поведения в соответствии с изменившимися условиями подкрепления (Richardson, 1973). Принимая этот вывод, необходимо уточнить, что сама миндалина относительно «безразлична» к вероятности подкрепления, к его информационному компоненту. Переход к 50 %-ному подкреплению вызывает фрустрацию у контрольных и септальных крыс, не оказывая влияния на амигдалэктомированных животных, в том числе на процесс угашения условных рефлексов, подкрепляемых в 50 % проб (Henke, 1977). М. Л. Пигарева (1978) вырабатывала пищевые условные двигательные рефлексы у крыс после двустороннего разрушения миндалин (рис. 11). Оказалось, что скорость выработки рефлексов у этих животных не отличается от аналогичных показателей у интактных крыс ни при высокой (100 и 50 %), ни при низкой (25 %) вероятности подкрепления. Только вероятность 33 %, для которой характерен максимум возникающего при этом эмоционального напряжения, сопровождается некоторым замедлением процесса формирования условного рефлекса (рис. 12).
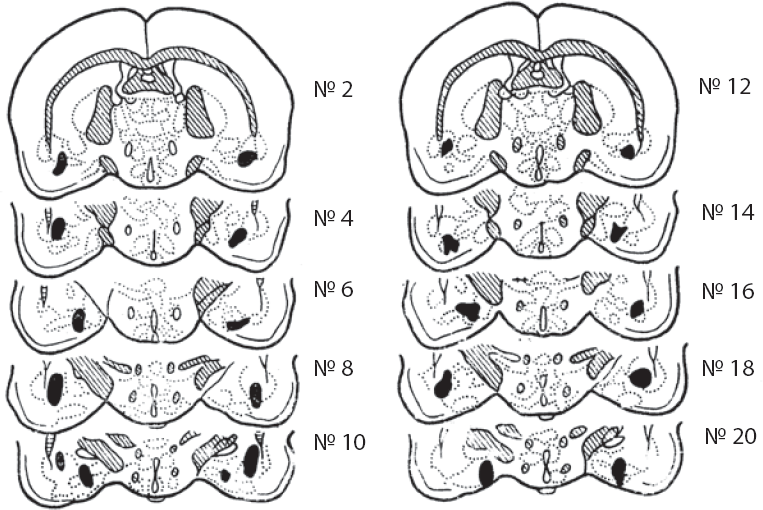
Рис. 11. Реконструктивная схема повреждения миндалевидного комплекса у 10 крыс

Рис. 12. Процент пищевых условных реакций у контрольных (1) и амигдалоэктомированных (2) крыс при различной вероятности подкрепления условного сигнала
Абсцисса – дни опытов (по М. Л. Пигаревой)
Двустороннее разрушение миндалины не препятствует выработке как пищевого, так и оборонительного условных рефлексов у крыс (рис. 13). По данным Р. Расселла и Р. Томпсона (Russell, Thompson, 1984), миндалина не играет ключевой роли в выработке мигательного условного рефлекса на тон у кроликов.