Книги
Мотивированный мозг. Высшая нервная деятельность и естественно-научные основы общей психологии
Итак, феномен самораздражения реализуется с участием двух основных медиаторных систем: норадренергической и дофаминергической. Можно предполагать, что норадреналин связан с побуждающими, мотивирующими компонентами этой реакции, а дофамин – с подкрепляющим, «награждающим» эффектом. Замечу, что, по мнению X. Маттиеса (Matties, 1982), подкрепляющая функция эмоций реализуется через дофаминергическую систему, обеспечивая долговременную память.
О преимущественном отношении норадреналина к механизмам мотивации свидетельствует и тот факт, что норадреналин возбуждает большую часть нейронов гипоталамуса у сытых крыс и тормозит у голодных (Dean, Dyer, 1984). Введение норадреналина в перифорникальную область передней части латерального гипоталамуса можно сделать подкрепляющим фактором для обучения сытых крыс предпочитать один из рукавов лабиринта. Дело в том, что норадреналин побуждал крыс есть, вторично активируя механизм положительных эмоций, связанных с актом еды, и делал привлекательными для животного пищевые стимулы (Cytawa, Jurkowlaniec, 1978; Cytawa, Jupkowlaniec, Bialowas, 1980). Интересно, что в отличие от гипоталамуса введение норадреналина в дорсомедиальную часть миндалины могло служить подкреплением для выработки условного рефлекса только у голодных крыс и теряло эти свойства у сытых (Jurkowlaniec, Bialowas, 1981). Мы склонны объяснять этот эффект следующим образом. При введении в миндалину норадреналин сохраняет свои побуждающие функции, но в отличие от гипоталамуса проявляет их только в отношении доминирующей, ранее актуализированной потребности, в данном случае голода.
Роль холинергической системы отчетливо проявляется в агрессивном поведении животных (Закусов, 1972), причем холинергический триггерный механизм агрессии испытывает на себе модулирующее влияние других медиаторных систем, в том числе норадреналина (Johansson, 1974; Romaniuk, Golebiewski, 1979). По мнению А. X. Алликметса (Allikmets, 1974), норадреналин ответствен за эффекторные проявления агрессии. Иными словами, он сохраняет свои побуждающие свойства и в отношении оборонительного поведения. При сочетании звука с ударами тока введение адреналина позволяет выработать условный рефлекс у крыс, находящихся под пентобарбиталовым наркозом (Weinberger, Gold, Sternberg, 1984). Что касается холинергической системы, то она преимущественно обеспечивает информационные компоненты поведения. Показано, что холинолитики влияют на пищедобывательные действия животного, на преодоление препятствий, на выбор пищи, но само состояние голода сохраняется (Вальдман, Козловская, 1972). Холинолитики нарушают совершенство и точность двигательных рефлексов избегания, не устраняя реакцию на боль.
К. Нолл и Дж. Дэвис (Noll, Davis, 1982) предлагают следующую функциональную классификацию нейромедиаторных систем. Возбуждающими медиаторами положительных эмоций («награды») являются катехоламины, отрицательных эмоций («наказания») – ацетилхолин. Серотонин играет роль тормозного медиатора в отношении обеих систем. К мнению об универсальном тормозящем влиянии серотонина на эмоциогенные структуры присоединяются и другие исследователи (Panksepp, 1982). Ослаблением тормозного влияния серотонина склонны объяснять повышенную агрессивность. Так, агрессивные мыши имеют более низкий уровень содержания серотонина в гипоталамусе, миндалине и гиппокампе по сравнению с неагрессивными (Serri, Ely, 1984). Содержание серотонина в среднем мозге и гипоталамусе ручных лисиц в 2 раза, а в гиппокампе в 1,5 раза выше, чем у диких (Попова, 1983). Введение предшественника дофамина повышает агрессивность крыс, а введение предшественника серотонина угнетает ее. Возможно, что первый эффект объясняется блокадой серотонинергической системы (Алликметс, Жарковский, 1976). Подавление серотонинергической системы усиливает половое поведение крыс, в то время как повреждение норадрен– и дофаминергической систем нарушает механизмы половой мотивации и удлиняет постэякуляторный период (McIntosh, Barfield, 1984). Р. Ш. Ибрагимов (1980) объясняет облегчение выработки условных пищевых рефлексов при введении серотонина в мозг его тормозным влиянием на механизмы ориентировочного рефлекса.
Представлениям об универсальном тормозном действии серотонина противоречат данные о его важной роли в процессах обучения на биологически отрицательном подкреплении. Введение р-хлорамфетамина нарушает выработку и сохранение реакций активного и пассивного избегания у крыс (Ogren, Johansson, Johansson, Archer, 1982). Исследования на моллюсках показали, что пластические изменения в центральной нервной системе связаны с увеличенным выходом серотонина из пресинаптического нейрона (Brunelli, Demontis, 1984). Серотонин участвует в ответах нейронов большого ядра шва мозга крыс на болевое раздражение (Dickenson, Goldsmith, 1984). П. М. Балабан с соавторами в опытах, поставленных в нашей лаборатории, показал, что дегенерация серотонинергических нейронов, вызванная введением 5,7-диокситрипта-мина, препятствует сенситизации безусловных оборонительных реакций виноградной улитки и выработке условного оборонительного рефлекса на пищевой раздражитель. С другой стороны, аппликация серотонина может быть использована в качестве подкрепления при его сочетании с пищевым стимулом. Возможно, что подкрепляющее действие серотонина связано с его влиянием на проводимость кальциевых каналов мембраны нервных клеток.
Можно предложить два объяснения противоречивым данным о роли серотонина в процессах обучения на эмоционально-отрицательном (болевом) подкреплении. Первое из них сводится к тому, что серотонинергические механизмы связаны не столько с выработкой условных рефлексов, сколько с их фиксацией, с процессами консолидации следов памяти независимо от эмоциональных характеристик подкрепляющего фактора (Кругликов, 1978). Второе объяснение состоит в следующем. Условные рефлексы на эмоционально-положительном подкреплении требуют специфического медиатора. Им является дофамин. Кроме того, здесь, по-видимому, участвуют энкефалинсодержащие нейроны (Stein, 1978; Cytawa, Trojniar, 1978). Лей-энкефалин облегчает реакцию самораздражения и угнетает реакцию избегания при стимуляции центрального серого вещества, а введение налоксона снижает частоту самораздражения эмоционально-позитивных структур. Что касается выработки оборонительных условных рефлексов, то здесь подкреплением служит
Таким образом, можно предложить следующую гипотетическую схему нейрохимической организации эмоций, опирающуюся на потребностно-информационную теорию. Информационные компоненты любой эмоции реализуются главным образом за счет холинергических структур. Для побуждающих, мотивационных компонентов наиболее существенна норадренергическая система. Что касается собственно эмоционального подкрепляющего звена, то в случае положительного подкрепления им является дофамин плюс эндогенные опиаты. При эмоционально-отрицательном подкреплении важную роль играет серотонин.
Данные, полученные на животных, хорошо согласуются с результатами клинических исследований. В опытах Дж. Эллисона (Ellison, 1975) снижение уровня норадреналина в мозге крыс делало их вялыми в знакомой обстановке и бесстрашными в новой, где новизна оказывалась мощным мотивирующим фактором. Снижение уровня серотонина давало обратный эффект: ослабление тормозного (стабилизирующего) влияния превращало новую ситуацию в чрезмерный, стрессирующий фактор, подавляя активность животных. Автор рассматривает эти два варианта в качестве моделей патологического состояния человека, где падение уровня серотонина порождает немотивированную тревогу, а дефицит норадреналина – депрессию. Действительно, мозг больных, покончивших с собой в состоянии депрессии, оказался обедненным как норадреналином, так и серотонином, причем дефицит норадреналина проявляется депрессией тоски, а дефицит серотонина – депрессией тревоги (Нуллер, 1981; Maas, Fawcett, Dekirmenjian, 1972; Sachar, Baron, 1979).
Для действия холинолитиков характерен психоз с преимущественным поражением интеллектуальных (информационных) процессов (Селиванова, Голиков, 1975).
Анализируя нейроанатомические и нейрохимические основы эмоций, мы убедились в том, что противоречивость имеющихся в настоящее время данных не удается преодолеть, рассматривая эмоции как автономный, замкнутый на себя феномен. Их изучение должно быть включено в более широкий контекст организации целенаправленного поведения, высшей нервной (психической) деятельности и еще шире – в контекст эволюции живой природы, а для человека – в процесс его культурно-исторического развития. Эта настоятельная необходимость и определила содержание следующей главы, где мы вынуждены вернуться к рассмотрению фактов, полученных преимущественно в опытах на животных.
Впрочем, такие переходы от экспериментов на животных к исследованиям с участием человека и обратно представляют одну из характернейших черт творческого метода И. П. Павлова.
Глава 4
Физиологические механизмы организации целенаправленного поведения
Поведением мы называем такую форму жизнедеятельности человека и животных, которая изменяет вероятность и продолжительность контакта с внешним объектом, способным удовлетворить имеющуюся у организма потребность. Прерывание или предотвращение вредоносного воздействия на организм, удовлетворяя потребность сохранения особи, ее потомства, вида в целом, представляет частный случай целенаправленного поведения.
Рассматривая индивидуальное поведение как момент, звено процесса самодвижения и саморазвития живой природы, а индивидуальную творческую деятельность человека как фрагмент эволюции культуры, мы должны указать на физиологические механизмы, определяющие, с одной стороны, активность поведения живых систем, а с другой – приуроченность этого поведения к реально существующим условиям окружающей внешней среды. Такими двумя механизмами являются, с нашей точки зрения, доминанта А. А. Ухтомского и формирование условных рефлексов по И. П. Павлову. В едином процессе организации поведения, приобретения новых навыков механизм доминанты обнаруживается в виде стадии генерализации условных рефлексов (Павлыгина, 1973; Досталек, 1980).
Феномен генерализации как стадии активного поиска жизненно важных объектов постоянно встречается в естественном поведении животных. Только что вылупившиеся цыплята клюют любые контрастирующие с фоном предметы, соразмерные с величиной их клюва. Позднее они обучаются клевать только те, которые могут служить кормом. Реакцию улыбки у ребенка вначале вызывает любой приблизившийся к нему человек, затем – только знакомое лицо. Если медвежонок теряет мать, он начинает ходить кругами, постепенно увеличивая их радиус, и в конце концов натыкается на след. Случаи спасения дельфинами людей и животных объясняются не их «разумом» и «альтруизмом», а генерализацией родительского инстинкта: самки дельфинов рожают в воде и первое время поддерживают детенышей на своих спинах.
Одна из характернейших черт доминанты – способность доминирующей в данный момент рефлекторной системы откликаться на самый широкий круг внешних стимулов, в том числе впервые встретившихся в жизни животного и человека. Вместе с тем возникающие при этом реакции отнюдь не хаотический перебор случайных проб и ошибок. Осуществляя поисковое поведение, животное использует действия, извлекаемые из прошлого опыта. Минимум два фактора канализируют и лимитируют доминантный поиск. Во-первых, это качество доминирующей потребности. Так, испытывающие жажду крысы становятся чувствительными к любым сигналам, связанным с водой (Schallert, 1982). Вторым лимитирующим фактором служат экологическая адекватность (или неадекватность) стимулов и ранее накопленный опыт. При создании у кролика доминантного очага методом поляризации мозговой коры постоянным током, по В. С. Русинову, соответствующую реакцию легче получить на шелест бумаги, чем на звучание тона. Если обезьяна убедилась, что палка для доставания приманки из глубокой щели оказалась слишком тонкой и короткой, она выбирает следующую палку толще и длиннее, но не наоборот. Ранее накопленный опыт ограничивает и канализирует поиск, а поиск в свою очередь обогащает и трансформирует нервные связи (ассоциации), хранящиеся в памяти.
Процесс трансформации доминанты в упроченный специализированный условный рефлекс мы попытались схематически изобразить на рис. 6. Под влиянием побуждающего безусловного стимула (например, голодового возбуждения) специфическая пищедобывательная реакция приобретает доминантный характер: ее может вызвать самый широкий круг внешних раздражителей. Но только тот из них, чье действие один или несколько раз совпадет во времени с приходом афферентации от подкрепляющего безусловного стимула (пищи), становится условным сигналом. Подчеркнем, что здесь непременно вовлекаются нервные механизмы эмоций, сопоставляющие эту афферентацию с уровнем пищевой возбудимости.
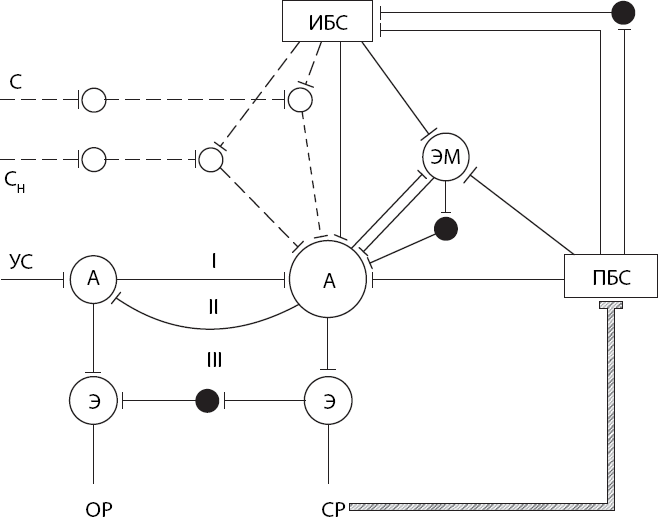
Рис. 6. Схема взаимодействия доминанты (прерывистые линии) и условного рефлекса (сплошные линии)
I – прямая условная связь; II – обратная активирующая условная связь; III – обратная тормозящая связь; черные кружки – тормозные элементы; тонкая линия – побуждающее влияние подкрепляющего стимула; заштрихованная линия – инструментальный условный рефлекс; С – Сн – индифферентные стимулы; УС – условный стимул; ИБС – инициирующий безусловный стимул; ПБС – подкрепляющий безусловный стимул; А – афферентные элементы рефлексов; Э – эфферентные элементы рефлексов; ОР – ориентировочная реакция; СР – специфическая реакция; ЭМ – эмоция