Книги
Мотивированный мозг. Высшая нервная деятельность и естественно-научные основы общей психологии
Именно наличие доминанты и ее устранение после действия условного сигнала (им может быть и собственная реакция животного) является условием быстрого (после одного, максимум двух сочетаний) образования условного рефлекса (Павлыгина, 1982; Павлыгина, Соколов, 1983). Напомним один из опытов Р. А. Павлыгиной (рис. 7).
После включения подпорогового постоянного тока, действующего в области коркового представительства левой лапы кролика, ранее индифферентные раздражители (тон, шелест) начинали вызывать движение этой лапы. Доминанта достигала своего оптимума через 20–25 мин после включения тока. В качестве показателя оптимального состояния в доминантном очаге для осуществления суммации учитывали наличие двигательной реакции конечности на три подряд примененных раздражителя и только после этого производили раздражение струей воздуха левого глаза животного. Если такая процедура вызывала не только мигание, но и движение соответствующей конечности, доминанту прекращали, выключая постоянный ток. Создание доминантного состояния спустя 20–30 мин в том же опыте приводило к произвольному миганию левым глазом, т. е. кролик совершал реакцию, направленную на прекращение доминантного состояния. Постоянный ток после этого выключали. Произвольное прекращение доминантного состояния закономерно наблюдалось в первом и во всех последующих опытах. Кролик мигал левым глазом через разные временные интервалы после включения постоянного тока. Это время измерялось минутами, а иногда секундами. Интервал между пробами был от двух до десяти минут. В первых опытах произвольное мигание иногда сопровождалось движением левой конечности. В последующих опытах движение конечности отсутствовало, что свидетельствовало о сужении рецептивного поля доминанты.
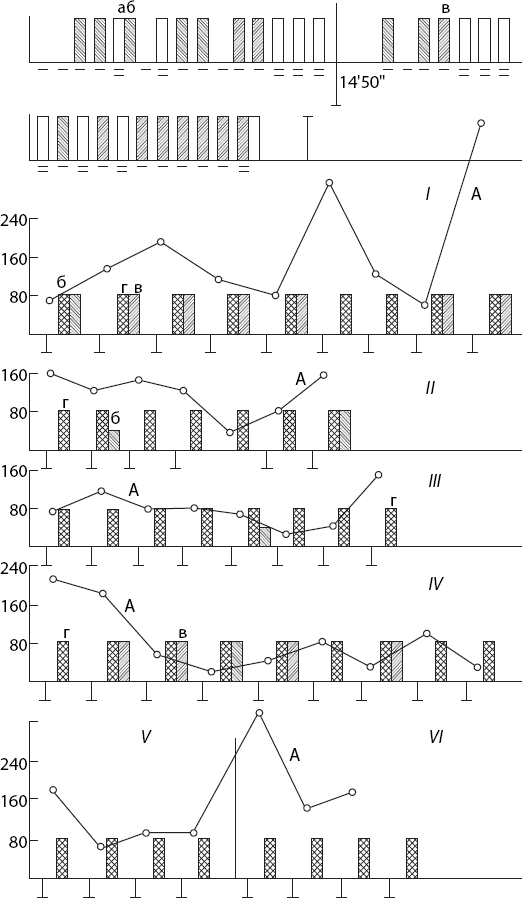
Рис. 7. Формирование целенаправленных реакций на основе стадии завершения доминанты
а – мигательный рефлекс на струю воздуха, направленную в глаз; б – общее движение животного; в – изолированное движение левой передней конечности; г – произвольное мигание; одна черточка – применение звукового раздражителя, две черточки – струи воздуха в глаз; вертикальная черта вниз – включение постоянного тока, вертикальная черта вверх – выключение постоянного тока; ордината – время (с); А – время появления произвольного мигания после включения постоянного тока;
Открытие Р. А. Павлыгиной имеет принципиальное значение для физиологии высшей нервной деятельности.
Экспериментально обнаруженный ею феномен может служить наиболее адекватной моделью
Уместно вспомнить, что существование потенциалов готовности рассматривается рядом ученых (в том числе Дж. Экклсом) как одно из оснований дуалистического взгляда на соотношение мозга и психики (Костюк, 1985). Исследования Р. А. Павлыгиной могут служить еще одним свидетельством нейрофизиологических основ произвольности, не требующей привлечения каких-либо понятий, лежащих за пределами нейрофизиологии.
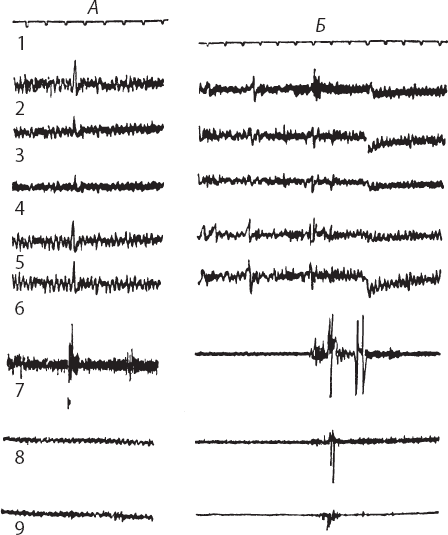
Рис. 8. Электрическая активность разных зон неокортекса при рефлекторном мигании, вызываемом струей воздуха, направленной в глаз. (
1 – время (с); 2 – ЭЭГ фронтальной зоны правого полушария; 3 – ЭЭГ сенсомоторной и 4 – теменной зон этого же полушария; 5 – ЭЭГ фронтальной и 6 – сенсомоторной зон левого полушария; 7 – активность мышц века; 8, 9 – активность мышц правой и левой конечностей (по Р. А. Павлыгиной)
Одновременная регистрация электрической активности мозговых структур, связанных с миганием, позволила установить определенную последовательность во времени перестройки потенциалов перед произвольным миганием. Раньше всего перестройка частотного спектра в сторону преобладания дельта-частот происходит в подкорковых центрах (за 8–4 с). Она более выражена по сравнению с перестройкой в коре перед самым миганием. Характерно, что преобладание дельта-частот в электрической активности подкорки сохраняется после выключения постоянного тока, в то время как в коре уже устанавливается доминирование тета-частоты. Можно думать, что вовлечение подкорковых образований мозга представляет существенный момент быстрого формирования условнорефлекторных связей после одного сочетания. По данным Р. Ю. Ильюченка (1981), для выработки условной реакции избегания после одного сочетания необходима сохранность миндалевидного комплекса и центрального серого вещества.
Условные рефлексы, формирующиеся после одного сочетания, встречаются достаточно часто. Так, пчела запоминает цвет кормушки после единственного посещения и сохраняет память о нем в течение 5–6 дней. Решающее значение для таких рефлексов имеют биологическая ценность (сила) подкрепления и экологическая адекватность стимула. Условное пищевое отвращение («эффект приманки»), когда подкрепляющее отравление возникает через несколько часов после действия условного сигнала, формируется в том случае, если этим сигналом служит вкусовой раздражитель, но не звук, освещение и т. п. Существует тенденция выделять рефлексы, замыкающиеся после одного сочетания, в особую разновидность высшей нервной деятельности, именуемую «психонервной» или «поведением, направляемым образами внешних объектов». Эксперименты Р. А. Павлыгиной побуждают тщательно проследить: в какой мере мы имеем дело с доминантным состоянием человека и животных во всех тех случаях, где наблюдается быстрое формирование новых условных связей? Целый ряд экспериментальных фактов, полученных другими авторами, делают это предположение весьма правдоподобным. Например, слабое болевое подкрепление не приводило к выработке реакции избегания у мышей и крыс. Предварительное воздействие болевым стимулом («сеанс страха») надолго, до 2 недель, облегчает последующее формирование условного рефлекса (Абуладзе, 1985). Предварительная навязанная стимуляция одного из позитивно-эмоциогенных образований мозга ускоряет выработку инструментальной реакции самораздражения этой или близкой к ней структуры (Орджоникидзе, 1985).
Наконец, исследования Р. А. Павлыгиной приближают нас к уяснению физиологических механизмов «закона эффекта» Торндайка, согласно которому однократное подкрепление пищей, прекращением болевого раздражения и т. д. фиксирует то движение животного, которое привело к достижению цели, хотя неэффективное движение повторялось до этого десятки и сотни раз.
Но вернемся к рассмотрению нашей схемы формирования условного рефлекса (см. рис. 6). После совпадения во времени действия условного раздражителя с приходом афферентации от подкрепляющего безусловного стимула начинает функционировать система обратных условных связей. При этом активирующая обратная связь избирательно повышает чувствительность структур, воспринимающих условный сигнал, в то время как тормозная устраняет эффекторные проявления «собственной» безусловной реакции на раздражитель, превратившийся в условный. Подкрепляющая афферентация тормозит влияние побуждающей (феномен «сенсорного насыщения»), но на ранних этапах удовлетворения соответствующей потребности она может усилить побуждение по принципу «аппетит во время еды». Наконец, зависимость подкрепления от реакций субъекта превращает классический условный рефлекс в его инструментальную разновидность.
Взаимодействие прямых и обратных условных связей Э. А. Асратян положил в основу своей рефлекторной теории мотивации поведения (Асратян, 1974). Это взаимодействие Э. А. Асратян рассматривал в качестве фундаментального общенейрофизиологического принципа интегративной деятельности мозга (Асратян, 1981). Об универсальности механизма двусторонних условных связей свидетельствует его наличие у беспозвоночных, что было показано при регистрации как поведенческих реакций (Максимова, 1979; Alexander, Audesirk, Audesirk, 1982), так и активности соответствующих нейронов (Максимова, Балабан, 1983; Балабан, Максимова, Галанина, 1985). Выработка пищевых условных рефлексов показала, что старые улитки способны обучаться только при наличии голода. Молодых улиток можно обучить и в сытом состоянии, но условный рефлекс проявляется у них лишь после пищевой депривации (Audesirk, Alexander, Audesirk, Moyer, 1982).
Экспериментальным подтверждением идеи Э. А. Асратяна о роли обратных условных связей в мотивации поведения может служить тот факт, что количество обратных связей уменьшается по мере насыщения животного, как это было показано В. Н. Семагиным (1983) на примере питьевых инструментальных реакций у крыс (рис. 9,
Хорошим методическим приемом для исследования роли обратных условных связей в организации поведения может служить анализ пространственной синхронизации электрической активности мозговых структур по М. Н. Ливанову (1972), в том числе при самостимуляции крыс слабым постоянным током (Павлыгина, Труш, Михайлова, Симонов, 1977). На рис. 10 показан процент случаев регистрации значимой когерентности (Р меньше 0,05) на частотах альфа– и тета-диапазонов при наличии соответствующей частоты в потенциалах сравниваемых мозговых образований: эмоционально-позитивный пункт гипоталамуса – двигательная область коры (1), эмоционально-негативный пункт гипоталамуса – двигательная область коры (2), двигательная кора, зрительная кора (3). Анализ проводился на следующих этапах поведения: I – спокойное состояние животного, II – непосредственно перед нажатием на педаль, III – во время пребывания на педали, IV – период ухода с педали, V – после ухода с педали. Рассматривая рисунок, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в 3 раза) возрастает процент случаев статистически значимой когерентности электрической активности эмоционально-позитивного пункта гипоталамуса с моторной областью коры. Возросшая когерентность свидетельствует о готовности нервных путей к проведению возбуждения по трем каналам обратных условных связей: от ранее раздражавшегося эмоционально-позитивного пункта к двигательной коре, к зрительной коре, а также между зрительным анализатором, воспринимающим условный сигнал будущего подкрепления (вид педали, ее местонахождение в камере и т. п.), и двигательной корой, поскольку именно вид педали будет направлять движение животного, инициированное следовым возбуждением эмоционально-позитивной зоны.
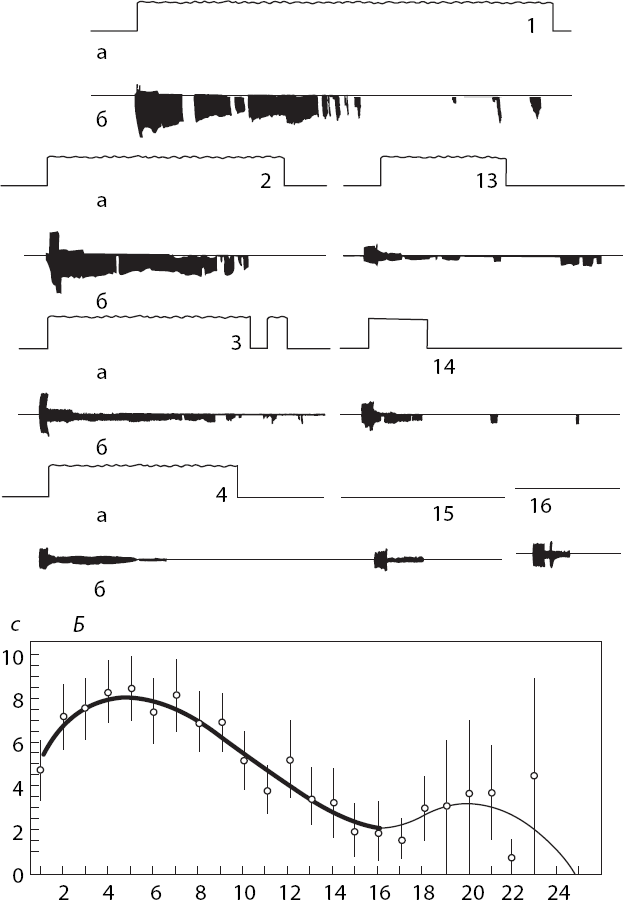
Рис. 9. Обратные условные связи при питьевом поведении крысы
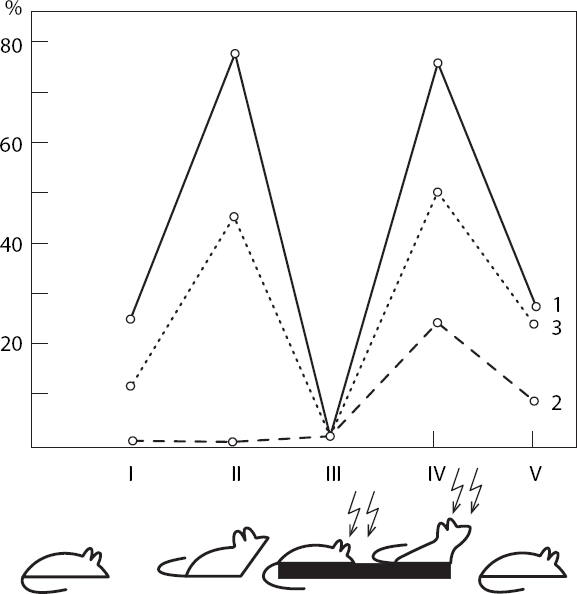
Рис. 10. Процент случаев статистически значимого изменения когерентности электрической активности мозговых структур на различных этапах самораздражения крыс
I – спокойное состояние: II – перед посадкой на педаль; III – на педали; IV – перед уходом с педали; V – сразу же после ухода; 1 – эмоционально-положительный пункт гипоталамуса – двигательная область неокортекса; 2 – эмоционально-отрицательный пункт – двигательная область неокортекса; 3 – двигательная область – зрительная область неокортекса (по Р. А. Павлыгиной, В. Д. Трушу, Н. Г. Михайловой, П. В. Симонову)