Книги
Огарок во тьме. Моя жизнь в науке
Когда после моей лекции задавали вопросы (которые благожелательно модерировал выдающийся биолог-теоретик Стюарт Коффман), кто-то в шутку спросил, может ли моя биоморфная программа выращивать не только алфавит, но и деньги. Я моментально вывел на экран вполне убедительный знак доллара (см. букву S в моей подписи на стр. 432), и так мой доклад завершился добродушным смехом.
Эмбриональный калейдоскоп
Хоть мой доклад в Лос-Аламосе и назывался “Эволюция способности к эволюции”, в тот момент я еще не развил эту тему на полную мощность. В главе “Эмбриональный калейдоскоп” в книге “Восхождение на гору Невероятности” я продвинулся в этой области дальше – в направлении, которое кажется мне весьма убедительным. Я уже упоминал о “зеркальных генах”, которые ввел в одной из поздних версий биоморфной программы. Можно представить, что гены, которые управляют симметрией животного в различных плоскостях, встраивают в эмбрион “зеркала”, подобные зеркалам в калейдоскопе. У большинства (но не у всех) животных такое зеркало стоит вдоль срединной линии: благодаря ему они двусторонне симметричны. Мутация в третьей ноге насекомого теоретически может затрагивать лишь правую сторону, но на практике отражается и на левой. Технически такое отражение ограничивает свободу эволюции: без него тоже можно было бы достичь – или, вернее, изловчиться построить – идеальную симметрию путем отдельных мутаций по двум сторонам (а еще так можно соорудить множество экзотических асимметрий впридачу). Но если предположить (основания для этого я приводил в книге “Восхождение на гору Невероятности”), что лево-правая симметрия сама по себе дает глобальные преимущества, то автоматическое отзеркаливание мутаций на обе стороны позволяет быстрее вносить эволюционные улучшения. Таким образом, необходимость симметрии (“зеркала” по срединной линии эмбриона-калейдоскопа) можно рассматривать не как ограничение (каким она, строго говоря, является), а, наоборот, как эволюционное усовершенствование способности к эволюции.
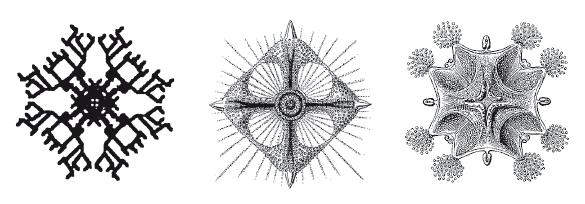
То же верно и для других плоскостей симметрии, хотя они реже встречаются в биологической реальности. На иллюстрации на следующей странице слева изображен компьютерный биоморф, обладающий четырехсторонней симметрией (два “калейдоскопических зеркала” под прямыми углами). В середине – строение радиолярии (изящного микроскопического одноклеточного существа), а справа – ставромедуза (конечно, в другом масштабе). У них всех есть “два зеркала” под прямыми углами, скрытые в эмбриологических глубинах. В случае биоморфа мне это доподлинно известно, потому что я сам написал его программу. О двух реальных животных я не знаю наверняка, но готов поставить что угодно, что их эмбриология по умолчанию ограничена четырехсторонней симметрией. Мое предположение заключается в том, что нововведение в фундаментальной эмбриологии, благодаря которому возникло это калейдоскопическое ограничение, давало определенные преимущества, и я бы назвал это нововведение эволюционным усовершенствованием способности к эволюции.
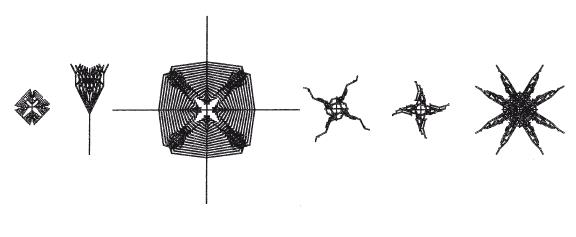
Иглокожие (морские звезды, морские ежи, змеехвостки и др.) в основном обладают пятилучевой симметрией. Опять же, мне кажется практически очевидным, что соответствующее правило симметрии скрывается в глубинах эмбриологии, и таким образом мелкая мутация, скажем, на кончике одной руки морской звезды, зеркалится на все пять рук (иногда рождаются морские звезды, у которых больше пяти рук, но это не опровергает мое обобщение). Опять же, с учетом того, что симметрия морской звезде чем-то полезна, отзеркаливание мутаций помогает “срезать дорогу” (по сравнению с отдельными мутациями в каждой руке по очереди) и внести изменения, не отступая от пятилучевой симметрии. Следовательно, это явление вполне заслуживает, чтобы его учитывали по категории “эволюции способности к эволюции”. И важно, что все мои попытки вывести пятисторонне-симметричных биоморфов на экране компьютера окончились провалом. Это практически очевидно. Пятилучевой симметрии можно добиться, только если кардинально переписать эмбриологическую схему – что снова возвращает нас к теме эволюции способности к эволюции. “Иглокожие” биоморфы, которых мне удалось вывести на экране, – сплошное надувательство (см. иллюстрацию на этой странице). С виду они напоминают плоского морского ежа, морскую лилию, морского ежа, змеехвостку и двух морских звезд соответственно – но ни один из них не обладает пятилучевой симметрией.
Во времена конференции в Лос-Аламосе цветной “Мак” еще не придумали. Когда я наконец им обзавелся, очевидным шагом в расширении генома биоморфов было добавить новый набор генов, задающих цвет. Тогда же я добавил гены, влияющие на линии, которыми были нарисованы первичные деревья эмбриологического алгоритма. Простые линии все еще дозволялись, но я ввел новый ген, меняющий их толщину, и другие гены, что превращали простые линии в прямоугольники или овалы, оставляли эти формы пустыми или закрашивали их, а также управляли цветом линий и заливки. Эти дополнительные гены открыли дорогу для новой мощной эволюции, искушая человека отбирать биоморфы, чтобы выводить подобия экзотических цветов, ковриков и бабочек. Мне вздумалось вынести компьютер в сад и предложить настоящим пчелам и бабочкам выбирать “цветы” и “бабочек” на экране. Я надеялся, что настоящие насекомые выведут из неопределенных форм подобия настоящих цветов. К сожалению, оказалось – как мне следовало предвидеть, – что яркий дневной свет, привлекающий насекомых, делает изображение на экране практически невидимым. И, как часто бывает с идеями, которые кажутся великолепными, я засунул ее в дальний угол и больше к ней не возвращался. Быть может, подойдут ночные мотыльки? Быть может, сенсорный экран, например, как у
Я работал над созданием цветных биоморфов, когда познакомился с Лаллой. Среди множества ее талантов – вышивка крестом (тогда она еще не увлеклась мозаикой, росписью керамики, ткачеством, а также вышивкой на машинке – причем последними двумя она занимается и ныне). Цветные, четырехсторонне-симметричные биоморфы вдохновили ее вышивать подушки и чехлы для стульев: стежки вышивки в точности соответствуют пикселям с экрана компьютера (см. цветную вклейку). Они вызывают восхищение и двадцать лет спустя.
Во всех моих биоморфных программах действовал искусственный, а не естественный отбор. Намного более трудная задача – в интересной форме сымитировать естественный отбор; об этом я мог лишь мечтать. Показательно уже само то, насколько это трудно. Можно представить, как встроить в биоморфную программу критерий отбора – скажем, по “колючести” или “округлости”. Ради эксперимента так я и сделал. Таким образом удалось обойти человеческий глаз как субъект отбора, и это сработало. Но это не представляло особого биологического интереса. Чтобы сымитировать выживание в мире, пришлось бы сконструировать этот мир, его физику, его (в идеале трехмерную) географию, его правила, по которым биоморфы взаимодействуют с другими объектами и с другими биоморфами, правила, по которым разные предметы не могут занимать одно и то же физическое пространство, и так далее. За годы, прошедшие с публикации “Слепого часовщика”, умные программисты разрабатывали подобные искусственные миры и их физику: например, Стив Грэнд и его
Артроморфы
В центре рассуждений об эволюции способности к эволюции – “шлюзы”, открывающиеся для новых творческих улучшений. Лос-Аламосская конференция, на которой я представил эту концепцию, сама по себе стала ее метафорой: на той конференции высвободилась некая творческая волна в моем собственном уме (и, вероятно, в умах других участников). Для меня эта волна достигла пика в “Восхождении на гору Невероятности”: среди моих книг эта кажется мне самой недооцененной (ее читают меньше всего – хотя, пожалуй, по новаторству она уступает только “Расширенному фенотипу”).
А вот еще одни ворота, открывшиеся на той конференции. Именно там я познакомился с Тедом Келером. Тед – один из блестящих программистов
У исходных биоморфов из “Слепого часовщика” было девять генов. У лос-аламосской версии – шестнадцать. У цветной – тридцать шесть. Каждое расширение генома открывало то, что я называл шлюзами, выпуская на свободу новое эволюционное “творчество”, хоть и с “конструктивными” ограничениями – например, сегментацией или “зеркалами калейдоскопа”. Но каждое из этих расширений было порождено вмешательством программиста. Мне каждый раз надо было начинать практически с нуля и дописывать массу нового кода. В каком-то смысле это служило уместной метафорой для эволюции способности к эволюции: думаю, что и в настоящей биологии переломные изменения, о которых мы говорим, – скажем, возникновение сегментации, многоклеточности, половых различий, пятисторонней симметрии у иглокожих – это редкие и катастрофические сдвиги, чем-то подобные кардинальному переписыванию компьютерной программы. Аналогию можно распространить даже на отладку программ. Ведь можно не сомневаться, что, когда в генофонд при отборе встраивается революционная мутация, от нее возникает множество побочных воздействий, которые постепенно сглаживаются: последующий отбор благоприятствует множеству дополнительных мелких мутаций, смягчающих побочные эффекты в целом полезной масштабной мутации.
Но настоящая биология знает и промежуточный уровень мутаций – не такой революционный, как возникновение многоклеточности, пола, сегментации или новых “зеркал симметрии”, но более масштабный, чем обычные точечные мутации, в которых один нуклеотид Уотсона – Крика заменяется на другой из четверки – С, Т, G или А. В этой промежуточной категории – дупликация (или ее противоположность, делеция) целых участков хромосомы. Дупликация – основной способ увеличения генома. В книге “Рассказ предка” (а именно, в рассказе миноги) я описал ее на примере гемоглобина. Вкратце: у нас есть пять разных типов белков-глобинов, которые кодируются разными генами в разных частях генома. А главное, что все пять произошли от одного-единственного предкового глобина, который кодировался единственным предковым геном. Предковый ген (у миног – наших отдаленных, примитивных родственников – до сих пор есть лишь он один) успешно дуплицировался в процессе эволюции и породил несколько “глобиновых генов”, которые мы имеем сегодня. Обычно, когда мы говорим об эволюционной дивергенции, мы имеем в виду расщепление исходного предкового вида надвое. Две популяции животных разделяются и идут разными дорогами. Здесь же мы все еще имеем дело с эволюционной дивергенцией, но говорим о расщеплениях, которые произошли
К слову, меня часто спрашивают: если бы я переписывал “Эгоистичный ген”, изменил бы я его содержание под влиянием нашего прогресса в понимании геномики? Ответ – “нет”, хоть местами и нерешительный: ученые ведь гордятся своей способностью менять точку зрения с появлением новых данных. Но мой “взгляд с точки зрения гена” с 1976 года, скорее, наоборот, упрочился благодаря новым данным, таким как дупликация генов, описанная в рассказе миноги. Теперь мы рассматриваем эволюционную дивергенцию на генном уровне, внутри отдельных особей, то есть нам становится виднее, что отбор скорее действует на генном уровне, а вовсе не на уровне индивидов.
Когда мы с Тедом разрабатывали характеристики артро-морфной программы, мы не пытались имитировать саму по себе дупликацию генов, как у гемоглобина. Тем не менее в нашей новой программе в некотором виде присутствовала генная дупликация (и делеция), из чего мы извлекли немало пользы. Во всех моих предыдущих биоморфных программах генный репертуар был закреплен (в разных версиях генов было девять, шестнадцать или тридцать шесть), у артроморфов же количество генов варьировало – оно само по себе было подвержено мутациям. Видите, как мы двигались к тому, чтобы позволить эволюции самой переписывать программу? Ведь прежде, каждый раз, когда способность биоморфов к эволюции возрастала благодаря макромутациям, это происходило потому, что я садился и писал гору нового кода.
Генетически артроморфы могли иметь и всего один сегмент, но сегментация вплеталась в самые основы их эмбриологии. По умолчанию действовало ограничение лево-правой зеркальной симметрии: она соблюдалась для всех артроморфов. Каждый сегмент состоял из овальной части тела (ее форму и размер задавали гены), способной отрастить пару симметричных конечностей, а каждая конечность могла раздвоиться в клешню. Пока все вполне членистоного. Число суставов в каждой конечности, размер и угол сочленения каждого сустава тоже определяли гены – как и размер и угол крепления клешней.
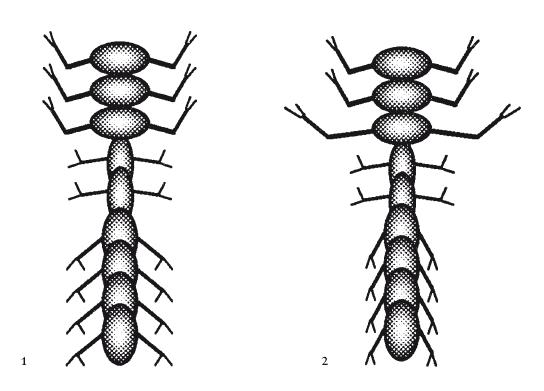
Эмбриологически любопытнее то, что группы соседних сегментов оказываются объединены в сферы влияния. Скажем, первые три сегмента могут быть практически неотличимы друг от друга, но заметно отличаться от двух следующих, и от следующих за теми четырех: такое строение напоминает голову, грудь и брюшко (см. Артроморфа 1 на иллюстрации ниже). Каждую группу сегментов (их не обязательно должно быть три – само это число тоже подвергалось генетическим мутациям) мы назвали “тагма”: этот греческий термин употребляется в биологии членистоногих. Но сегменты внутри тагмы не обязательно должны были быть полностью идентичными. На каждый сегмент влияли его собственные тагмаспецифические гены, которые могли мутировать независимо от других сегментов. Сравнительное единообразие внутри тагмы достигалось умножением численных генетических параметров каждого сегмента на определенный коэффициент (“ген”), присущий конкретной тагме. Изображенный здесь Артроморф 2 похож на Артроморфа 1, за исключением того, что сегмент 3, хоть очевидно и принадлежит к тагме 1, обладает более длинными ногами, чем два других сегмента в тагме 1. Также в тагме 3 мы в целом наблюдаем совсем другие ноги.
На более высоком уровне присутствовали и другие гены, на которые умножались параметры всех генов всего организма, во всех тагмах. И наконец, мы добавили гены градиента, которые умножали другие генетические воздействия на некий растущий (или убывающий) коэффициент по мере продвижения к задней части организма (или тагмы). Количество тагм и сегментов внутри каждой тагмы увеличивалось (или уменьшалось) посредством дупликации (или делеции) генов.
Такова была эмбриология артроморфов, и вы заметите, что она оказалась сложнее, чем эмбриология биоморфов – в биологически интересных смыслах. Я работал на пределе своего умения программировать и вынужден был полагаться на мастерство Теда, который намного меня превосходил. Код я писал самостоятельно (на “Паскале” – не самом любимом языке Теда, а теперь и вовсе давно устаревшем), но Тед направлял меня указаниями по электронной почте на псевдокомпьютерном языке, напоминающем формальную сокращенную версию английского. Иногда, подозреваю, моя неповоротливость приводила его в легкое раздражение – я точно не дотягивал до уровня профессиональных программистов

Череда конференций Криса Лэнгтона “Искусственная жизнь” продолжалась, как и “Цифровая биота”, другая похожая серия. Сам Крис присутствовал на второй конференции в кембриджском Модлин-колледже в 1996 году; меня пригласили прочесть на ней основной доклад под названием “Взгляд с точки зрения реальной жизни”: то была очевидная попытка “укоренить” компьютерщиков, увлеченно исследующих виртуальные миры, в настоящей биологии. Мне конференция запомнилась главным образом благодаря чудесному экспромту Дугласа Адамса (текст его выступления приведен в книге “Лосось сомнений”) и знакомству со Стивом Грэндом – автором книги “Сотворение: жизнь и как ее создать”, шедевра, равного по масштабам его программе искусственной жизни,