Книги
Эмоциональный мозг
Тесная связь функций гипоталамуса и миндалины продемонстрирована множеством экспериментов. По данным Т. Н. Ониани, эффекты стимуляции миндалины должны быть опосредованы через гипоталамус и другие структуры мезодиэнцефалона [Ониани, Мгалоблишвили, Чиджавадзе, 1978]. Миндалина оказывает регулирующее влияние на функции гипоталамуса, а в случае их выпадения компенсирует образовавшийся дефект. Эта компенсация происходит с участием структур новой коры. Так, после компенсации афагии у крыс, вызванной повреждением латерального гипоталамуса, корковая распространяющаяся депрессия вновь вызывает афагию, которая сохраняется в течение многих дней после декортикации. Повреждение ядер самой миндалины или полная нервная изоляция миндалевидного комплекса сопровождается менее выраженными поведенческими изменениями, чем аналогичные вмешательства на гипоталамусе.
В настоящее время имеется достаточно оснований рассматривать гипоталамус как сравнительно высокий уровень интегративной деятельности мозга. Очень трудно найти в гипоталамусе структуры, стимуляция которых вызвала бы вегетативные сдвиги без эмоциональных реакций. По-видимому, в гипоталамусе нет чисто «вегетативных центров», гипоталамус связан с организацией целостных поведенческих актов, в том числе — их вегетативных компонентов [Поляков, Талан, Черниговский, 1978]. Нейроны гипоталамуса очень быстро вовлекаются в условнорефлекторный ответ: активность нейронов латерального гипоталамуса изменялась через 150–200 мс после открывания заслонки для подачи пищи. Аналогичный ответ нейронов бледного шара наблюдался только через 300 мс, когда возникал двигательный компонент — изменения электромиограммы, связанные с лизанием [Rolls, Roper-Hall, Sanghera, 1977].
Реакция нейронов латерального гипоталамуса на вид и запах пищи зарегистрирована только у голодных обезьян, введение глюкозы устраняло этот эффект [Burton, Rolls, Mora, 1976]. Пачкообразная активность нейронов вентромедиального ядра гипоталамуса, возникающая у голодных животных, усиливается при поступлении пищи в рот и исчезает по мере наполнения желудка [Судаков, Журавлев, 1979]. Вместе с тем в гипоталамусе имеются и такие нейроны, которые будучи активированы голодом тормозятся сразу же с началом еды [Олдс, 1977]. При экспериментальном морфинизме у крыс нейроны латерального гипоталамуса активируются состоянием абстиненции (потребность) и тормозятся морфином (подкрепление). Нейроны медиального гипоталамуса, напротив, активируются подкреплением и тормозятся по мере актуализации потребности. Аналогичные данные получены для жажды и питья.
Многие авторы отмечают сравнительно узкую специализацию структур гипоталамуса. По данным Б. Оливьера, передний отдел медиального гипоталамуса у крыс контролирует пассивно-оборонительные реакции [Olivier, 1977]. Повреждение медиального гипоталамуса усиливает агрессивность крыс, вызванную болевым раздражением лап, и не влияет на территориальную агрессивность, которая нарушается только при разрушении латерального гипоталамуса [Adams, 1971]. Однако не меньшее количество фактов свидетельствует о зависимости последствий стимуляции и повреждения гипоталамуса от доминантного состояния животного и от стимулов окружающей среды [Isaacson, 1974].
Эффект стимуляции гипоталамуса одними и теми же параметрами тока при одной и той же локализации кончика электрода зависит от того, производится ли раздражение в пищевой или оборонительной ситуации опыта, у голодного или сытого животного [Беленков, Шалковская, 1978]. Стимуляция в области латерального гипоталамуса вызывает реакцию атаки на подчиненную крысу и не вызывает агрессивности по отношению к высокоранговому животному или самке [Koolhaas, 1978]. Под влиянием раздражения гипоталамуса макаки резусы нападают преимущественно на подчиненных самцов [Alexander, Parachio, 1973].
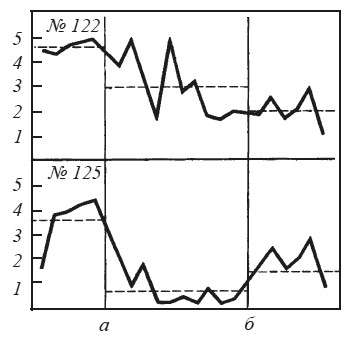
Для нас представляют особый интерес те случаи, где повреждение гипоталамуса дает эффект, противоположный амигдалэктомии. Так, после разрушений в гипоталамусе животные перестают реагировать на «тканевую жажду» и падение глюкозы в крови, но продолжают отвечать на условные сигналы воды и пищи [Олдс, 1977]. Их прошлый опыт приобретает известную самостоятельность, будучи изолирован от текущих нужд организма. Напомним, что при повреждениях миндалины нередко наблюдается прямо противоположное явление: например, после разрушения вентральной части медиального ядра у крыс нарушаются условные рефлексы, хотя потребление воды и пищи не претерпевает особых изменений [Korczynski, Fonberg, 1976]. Результат, противоположный последствиям амигдалэктомии, мы наблюдали и в опытах с реакцией избегания у крыс при болевом раздражении другой особи (рис. 35). Если амигдалэктомии нарушает баланс между конкурирующими мотивациями и выявляет доминирующую из них, то двустороннее повреждение латериального гипоталамуса, напротив, выравнивает силу мотиваций. Для этих крыс характерно «застревание» между педалью, включавшей ток, и открытым пространством камеры.
Итак, на уровне гипоталамуса конфликт между конкурирующими мотивациями однозначно решается в пользу одной из них за счет преобладания доминирующей в данный момент потребности. Участие миндалины делает этот процесс более пластичным, поскольку в конфликт вовлекаются эмоции, зависящие не только от силы потребностей, но и от вероятности их удовлетворения с учетом прошлого опыта и наличной ситуации. Благодаря миндалине возникает возможность сосуществования мотивов, их динамической иерархии. Эта возможность имеет огромное приспособительное значение: представим себе голодное животное, которое, стремясь к пище, прекращает наблюдение за сигналами потенциальной опасности. Вместе с тем функционирование миндалины способно осложнить поведение затрудненностью выбора между конкурирующими побуждениями. Подобно гиппокампу миндалину также можно назвать «органом колебаний и сомнений», но не в информационном, а в мотивационном смысле. Гиппокамп причастен к колебаниям, связанным с достижимостью целей, миндалина «взвешивает» их ценность.
Взаимодействие мозговых структур, судя по пространственной организации их биоэлектрических процессов
Спектрально-корреляционный анализ электрической активности различных образований головного мозга значительно пополнил сведения о функциональной организации ряда форм высшей нервной деятельности животных и человека. Наиболее обстоятельно были изучены пространственные отношения процессов, протекающих в различных мозговых структурах при ориентировочной реакции, при выработке и угашении условных рефлексов, в естественном и медикаментозном сне, при некоторых умственных операциях, совершаемых здоровым и больным человеком [Ливанов, 1972]. Значительно меньше известно о пространственной организации биоэлектрических процессов при возникновении состояний голода, жажды, полового влечения и во время поведения, направленного на удовлетворение этих биологических потребностей.
Хорошей моделью для изучения мотивированного поведения и его электрофизиологических коррелятов является феномен самораздражения. Однако традиционный способ самостимуляции ритмическим током исключает возможность изучения электрической активности раздражаемых структур, хотя возбуждение именно этих структур инициирует все последующие действия животного. Вот почему использование постоянного тока для самостимуляции животных впервые открыло принципиально новые возможности электрофизиологического исследования этой формы мотивированного поведения [Павлыгина, Михайлова, Симонов, 1975].
Систематическими работами В. С. Русинова [1969] и его сотрудников были показаны преимущества постоянного тока для создания стойких доминантных очагов в центральной нервной системе. Поляризуя гипоталамическую область головного мозга кролика, Р. А. Павлыгина [1956, 1958] получила доминантный очаг, оказывающий выраженное влияние на оборонительные условные рефлексы, электрическую активность различных мозговых образований и кровяное давление. Приступая к экспериментам, мы не нашли в литературе работ, посвященных поляризации эмоциональных «центров» и феномену самораздражения постоянным током, равно как и данных о пространственной организации электрической активности определенных образований мозга при мотивированном поведении животных. Совместно с Р. А. Павлыгиной, В. Д. Трушом и Н. Г. Михайловой опыты были поставлены нами на 16 белых крысах-самцах. У 6 из них на 6-канальном магнитофоне производилась регистрация суммарной электрической активности моторной и зрительной областей коры, гиппокампа, обонятельных луковиц, эмоционально-позитивных и негативных зон гипоталамуса (крысы № 13, 14, 15, 197, 221, 222).
Для монополярного раздражения использовали электроды с диаметром кончика 60 мкм, зачищенным от лака на 0,3–0,5 мм. Индифферентный электрод находился в мышцах шеи. Для получения феномена самостимуляции раздражали структуры медиального пучка переднего мозга в преоптической области и латеральном гипоталамусе. Эмоционально негативные реакции получали при раздражении дорзомедиальной области покрышки. Прежде чем поляризовать позитивные зоны, эти точки тестировались на эффективность в отношении самораздражения ритмическим током. С этой целью использовали однофазные, прямоугольные электрические импульсы частотой 1000 циклов в секунду при длительности стимулов 0,1 мс, длительности пачки 0,3–0,5 с интенсивностью 240–560 мкА. Поляризация малыми токами 2–6 мкА, которая обычно используется для создания доминантных очагов при экспозиции около одного часа, была неприемлема в наших экспериментах. Мы использовали токи 20–100 мкА, и животное не находилось на педали дольше 3 мин.
Спустя 10 дней после операции оживления электродов крысу помещали в плексигласовый ящик размером 21 × 25 × 40 см, у одной из стенок которого находилась педаль. Как правило, на протяжении первых 2–3 дней регистрировали количество нажатий на педаль, совершаемых крысой в процессе ее ориентировочно-исследовательской активности. Затем замыкали цепь, в результате чего каждое нажатие на педаль начинало сопровождаться воздействием анода или катода постоянного тока. Поляризации подвергали только те пункты, подкрепляющие свойства которых были предварительно установлены стимуляцией ритмическим током. Если крыса долго не вставала на педаль, ее сажали на педаль, удерживая в этом положении несколько секунд. Об эффективности поляризации эмоционально-позитивных зон гипоталамуса можно было судить по числу нажатий и продолжительности пребывания на педали, а также — по динамике угашения реакции самостимуляции после выключения тока. В ряде случаев влияние постоянного тока контролировалось по избеганию нажатий на педаль после поляризации эмоционально-негативных пунктов «наказания».
Четырехсекундные интервалы записи электрической активности мозговых образований обрабатывали на ЭЦВМ. При этом оценивали автоспектры потенциалов каждого исследуемого образования, а также кросскорреляционные коэффициенты и когерентные функции между потенциалами каждой пары изучаемых структур. Все спектральные характеристики оценивали в диапазоне от 1 до 20 Гц. Методика определения частотного спектра и вычисления функции когерентности описаны ранее [Труш, Кориневский, 1978]. Морфологический контроль мозга крыс подтвердил локализацию отводящих и стимулирующих электродов.
У крыс № 197, 221 и 222, у которых регистрировалась электрическая активность двигательной и зрительной областей новой коры, эмоционально-позитивной и негативной зон гипоталамуса, в фоновых потенциалах перечисленных структур согласно автоспектральным оценкам, как правило (примерно 80 % наблюдений), доминирующими были частоты тета- и альфа-диапазонов. Во время пребывания крысы на педали при действии подкрепляющего постоянного тока в 40 % записей наблюдалось доминирование дельта-волн. Приблизительно в 50 % случаев, когда наиболее выраженными оставались колебания тета- и альфа-диапазонов, частота доминирующего ритма уменьшалась на 1,5–3 Гц по сравнению с фоновыми записями. После ухода крысы с педали во всех без исключения случаях частота доминирующего ритма возрастала в 60 % — на 0,5–2 Гц.
При анализе оценок когерентных функций гипоталамических и корковых структур прежде всего обращает на себя внимание то, что каждому из изучавшихся в эксперименте этапов поведения соответствует определенный «рисунок» распределения корреляционных связей (рис. 36). Это подтверждает мнение М. Н. Ливанова [1972] о том, что именно в пространственной организации биопотенциалов находят отражение различные функциональные состояния мозга, что было убедительно показано, например, для ориентировочного рефлекса, для различных стадий выработки условного рефлекса и для некоторых тормозных состояний.
В табл. 5, где представлены результаты анализа оценок когерентных функций потенциалов мозга крыс № 221 и 222, приняты следующие обозначения. Цифры показывают процент случаев регистрации значимой когерентности (уровень значимости Р < 0,05) на частотах тета- и альфа-диапазонов при наличии соответствующих ритмических составляющих в потенциалах сравниваемых образований.
Рассматривая таблицу, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в три раза) возрастает когерентность электрической активности эмоционально-позитивного пункта гипоталамуса с ЭЭГ моторной и зрительной коры и когерентность потенциалов двух корковых участков. В то же время корреляционные отношения между потенциалами этих структур и эмоционально-негативной зоны гипоталамуса существенно не изменяются.
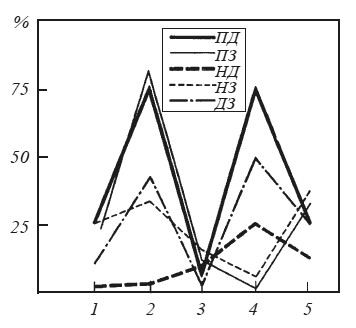
Рис. 36. Оценки вероятности наличия значимой когерентности между потенциалами мозговых структур на различных этапах поведения при осуществлении реакции самополяризации