Книги
Эмоциональный мозг
Для более объективного анализа изменений вегетативных функций в процессе выработки условнорефлекторного переключения на 5 гиппокампэктомированных и 4 интактных крысах была поставлена специальная серия опытов с регистрацией электрокардиограммы. В камере, разделенной перегородкой на два идентичных отделения, вырабатывали переключение разнородных (пищевого и оборонительного) условных рефлексов на один и тот же звуковой раздражитель. К выработке оборонительного условного рефлекса (двустороннего избегания) приступали после закрепления в течение 10–15 опытов пищевого условного рефлекса (открывание дверцы кормушки). Переключателем служил мелькающий свет, включаемый за 1 мин до первого предъявления звукового сигнала, сопровождавшегося болевым раздражением. Мелькающий свет действовал на протяжении всей «оборонительной» части опыта. В первой половине опыта 10 предъявлений звука (с интервалом 1–3 мин) подкрепляли пищей, после включения мелькающего света 10 предъявлений звукового раздражителя подкрепляли током (0,8–1,0 мА), подаваемым на проволочный пол, если крыса в течение 5 с не перебегала в соседнее отделение камеры.
Электрокардиограмму, двигательные реакции животного и отметки предъявляемых раздражителей регистрировали на самопишущем приборе. Для отведения электрокардиограммы за несколько дней до начала экспериментов всем крысам были вживлены специальные серебряные электроды, которые выводились в разрез, сделанный на бедре, и крепились к мышце. Подпаянная к электроду изолированная покрытием проволочка выводилась под кожей с помощью зонда к голове, где вместе с индифферентным электродом укреплялась на поверхности черепа.
В процессе выработки пищевого условного рефлекса общее поведение и условнорефлекторная деятельность интактных и оперированных крыс существенно не отличались. Частота сердечных сокращений у гнппокампэктомнрованных крыс, в среднем, была несколько ниже, чем у интактных, и составляла соответственно 345 и 385 ударов в минуту.
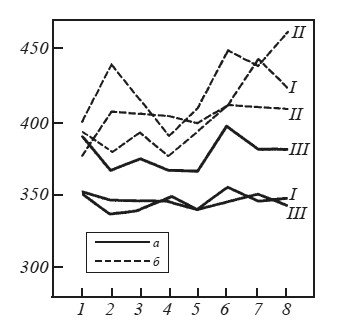
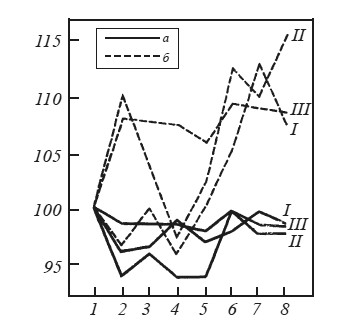
При введении оборонительного подкрепления у интактных крыс наряду с описанными выше вегетативными реакциями: интенсивной дефекацией во время предъявления звукового раздражителя и в межсигнальных интервалах, мочеиспусканием, пилоэрекцией, учащением дыхания значительно возрастала частота сердечных сокращений. На рис. 32 приводятся средние данные, отражающие динамику частоты сердечных сокращений при предъявлении раздражителей и при выполнении двигательных реакций. Видно, что предъявление звукового условного сигнала в первом опыте вызывает у интактных крыс снижение частоты сердечных сокращений, тогда как предъявление мелькающего света, как и осуществление пищевой двигательной реакции, возвращает этот показатель к исходному уровню. После болевого раздражения наблюдается сначала резкий подъем, а потом некоторое снижение частоты сердечных сокращений. Более наглядно эти данные представлены на рис. 33, где частота сердечных сокращений выражена в процентах по отношению к фону, принятому за 100 %. Видно, что резкое увеличение этого показателя происходит после электроболевого раздражения (через 2 и 10 с после выключения тока).
У гиппокампэктомированных крыс во время первого опыта по выработке переключения частота сердечных сокращений изменялась незначительно (рис. 32). В пищевой ситуации звуковой сигнал, также как у интактных крыс, вызывал снижение частоты сердечных сокращений. Выключение звука возвращало частоту сердечных сокращений к исходному уровню. Частота сердцебиений у оперированных крыс снижалась и при включении мелькающего света, а в момент действия тока и на протяжении первых 10 с после его выключения частота сердечных сокращений практически не изменялась. Таким образом, в первый день выработки переключения частота сердечных сокращений у гиппокампэктомированных крыс не отличалась от фона.
Во время второго опытного дня предъявление условного звукового раздражителя вызывало у интактных крыс не снижение, как в предыдущем опыте, а резкое увеличение частоты сердечных сокращений (см. рис. 32). После выключения звука она столь же резко падала до фонового уровня. Увеличение частоты сердечных сокращений наблюдалось и при предъявлении звука на фоне мелькающего света. Еще большее учащение происходило через 2 и 10 с после выключения болевого раздражения.
У гиппокампэктомированных крыс во время второго опыта частота сердечных сокращений также увеличивалась, особенно при предъявлении звукового раздражителя на фоне мелькающего света, однако резкие спады и подъемы кривой, характерные для интактных крыс, у них отсутствуют (см. рис. 32). Интересно, что в ответ на звуковой сигнал в пищевой ситуации оперированные крысы в отличие от интактных отвечают снижением частоты сердечных сокращений, а не резким учащением по сравнению с фоном.
На протяжении всего третьего опыта по выработке переключения как в пищевой, так и в оборонительной ситуации у интактных животных наблюдалась тахикардия, выраженная в большей или меньшей степени в зависимости от действующих раздражителей. Ярко проявлялись и другие вегетативные компоненты оборонительной реакции, причем адекватная двигательная оборонительная реакция у этих крыс, как правило, отсутствовала. Частота сердечных сокращений у гиппокампэктомированных крыс практически не отклонялась от фонового уровня (рис. 33). Во время третьего опыта у 4 оперированных крыс зарегистрировано уже 100 %-ное, а у одной — 90 %-ное адекватное выполнение пищевого и оборонительного условных рефлексов.
Теперь мы можем ответить на вопрос о том, относится ли гиппокамп к системе мозговых структур, формирующих эмоциональное напряжение. Гиппокамп
Гиппокамп
По-видимому, гиппокамп играет роль не только входного фильтра информации, подлежащей или не подлежащей регистрации в долговременной памяти [Виноградова, 1975], но принимает участие и в извлечении следов из памяти на предмет их использования в текущем поведении [Hirsh, 1974]. Если животное без гиппокампа целиком зависит от наличных стимулов и реагирует по принципу «стимул — реакция» (вспомним гиппокампэктомированных крыс в опытах с условнорефлекторным переключением), то в интактном мозге следы могут быть извлечены из памяти независимо от внешних стимулов и обеспечить ожидание этих стимулов по механизму тонических обратных условных связей [Асратян, 1974]. Фактором, активирующим энграммы ранее воспринятых раздражителей, в последнем случае служит возбуждение мозгового субстрата потребностей — голода, жажды и т. п. Возрастание количества реакций на сигналы маловероятных событий зависит от деятельности двух механизмов: от расширения диапазона актуализированных следов и от снижения критериев «принятия решения» при сопоставлении этих следов с наличными стимулами, поступающими из внешней среды.
Электрофизиологическим коррелятом механизма, квантующего поток извлекаемых из памяти энграмм, является тета-ритм, столь характерный для электрической активности гиппокампа. Все ситуации, в которых мы наблюдаем усиление тета-ритма, будь то ориентировочный рефлекс, поисковое поведение, организация сложных неавтоматизированных движений, появление признаков эмоционального напряжения и т. п., обладают одной общей для них чертой: перечисленные случаи требуют активной мобилизации ранее выработанных условных связей, извлечения хранящихся в памяти энграмм для сопоставления с поступающими извне сигналами или для «пересмотра», рекомбинации следов в целях построения новых приспособительных действий. Л. А. Преображенская [1978] показала, что повышение частоты и регулярности тета-ритма наблюдается в опытах с переключением у собак, когда: 1) животное недостаточно четко различает оборонительную и пищевую ситуацию; 2) не завершено формирование полноценного инструментального оборонительного рефлекса.
В результате своих систематических экспериментов Т. Н. Ониани пришел к заключению о том, что тета-ритм в гиппокампе свидетельствует о наличии мотивационного возбуждения, проявляющегося в поиске объектов, способных удовлетворить имеющуюся потребность. Оба этих симптома (тета-ритм и мотивированное поведение) можно наблюдать при стимуляции латерального гипоталамуса. Если стимуляция гипоталамуса не вызывает мотивированного поведения (например, при раздражении дорсомедиального ядра), в гиппокампе регистрируется десинхронизация электрической активности. Торможение мотивированного поведения (например, при стимуляции вентромедиального ядра) также сопровождается десинхронизацией в гиппокампе [Ониани, Унгиадзе, Абзианидзе, 1970]. Амплитуда и частота гиппокампального тета-ритма у крыс, голодавших 22 часа, заметно возрастают по сравнению с тета-ритмом крыс, подвергнутых 3-часовой депривации. Голодные крысы активно исследуют окружающую среду в отличие от малоподвижных сытых животных [Ford, Bremner, Richie, 1970].
Видовые особенности тета-ритма отнюдь не свидетельствуют о том, что у разных видов тета-ритм имеет совершенно различное функциональное значение, поскольку видовые характеристики тета-ритма коррелируют с особенностями ориентировочно-исследовательского поведения у представителей данного вида [Bennett, French, Burnett, 1978]. Несколько лет назад мы писали вместе с М. В. Фроловым: «Тета-ритм особенно характерен для ситуации активного поиска путей удовлетворения возникшей потребности» [Симонов, Фролов, 1970, с. 154]. Связь тета-ритма с внешним поисковым поведением, равно как и с пересмотром хранящихся в памяти энграмм (в том числе — в состоянии парадоксального сна), все больше привлекает внимание исследователей [Арашвский, Ротенберг, 1978].
Связь гиппокампального тета-ритма с процессами фиксации следов в памяти и с процессами извлечения этих следов обнаружена в экспериментах различного типа. На ранних стадиях выработки условного рефлекса (фиксация следов) тета-волны в гиппокампе опережают тета-волны в височной коре. При упрочении условной связи тета-волны энторинальной коры опережают волны в гиппокампе, происходит воспроизведение следов [Adey, Dunlop, Hendrix, 1960]. В опытах с опознанием определенных объектов высокая степень когерентности в пределах узкой полосы частот около 6 Гц между электрической активностью гиппокампа, зрительной и сенсомоторной коры сопровождала только правильные ответы [Эйди, 1969]. При однократном обучении крыс ударом электрического тока степень сохранения условного рефлекса хорошо коррелирует с выраженностью гиппокампального тета-ритма в период после обучения [Landfield, Me Gaugh, Tusa, 1972]. Стимуляция перегородки у кроликов и крыс с параметрами раздражения, которое усиливает тета-ритм в гиппокампе, ускоряла выработку условного рефлекса и способствовала его сохранению [Wetzel, Ott, Matthies, 1977; Кориневская, 1978]. Перечисленные и подобные им факты служат дополнительным свидетельством в пользу гипотезы о связи тета-ритма с механизмами извлечения следов из памяти и их сопоставления с наличными стимулами.
Процесс выработки условных рефлексов у интактных и гиппокампэктомированных животных имеет одно интересное отличие (см. рис. 31). При вероятности подкрепления порядка 100-50 % интактное животное сравнительно быстро начинает давать высокий процент условных реакций. У гиппокампэктомированных крыс в первые дни сочетаний условные реакции практически отсутствуют, а затем стремительно, на протяжении одного-двух опытных дней достигают критерия выработки. Создается впечатление, что в отсутствие гиппокампа продолжает работать какой-то «счетчик подкреплений» (по-видимому, новая кора), реализующий условные реакции только после набора статистики, надежно прогнозирующей подкрепление. Что касается мозга с неповрежденным гиппокампом, то для него достаточно сравнительно небольшого количества сочетаний, чтобы «выдвинуть гипотезу» о закономерном следовании подкрепления после каждого предъявления условного сигнала. К. Прибрам и Р. Дуглас считают одной из важных функций лимбических структур способность «сохранять предположение», несмотря на отвлечение внимания и неподкрепление [Pribram, Douglas, Pribram, 1969].