Книги
Огарок во тьме. Моя жизнь в науке
К этому вопросу можно подойти, например, проведя мысленный эксперимент, который в книге “Эгоистичный ген” я назвал “зеленой бородой”. Большинство новых мутаций оказывают несколько разных воздействий на тело (этот эффект называется плейотропией). Представьте себе ген, который дает особям приметную черту вроде зеленой бороды, а заодно наделяет добрыми чувствами к зеленым бородам и склонностью помогать зеленобородым индивидам выживать и размножаться. Как выразился бы Дживс, “этот вариант не выглядит осуществимым, сэр” – но суть этот образ передает. И подобный ген смог бы распространиться в популяции. Этот пример обрел популярность (поиск в
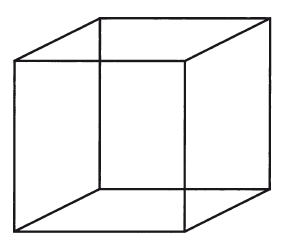
В 1964 году Гамильтон описал математический способ переопределить “приспособленность” таким образом, чтобы учитывать родственные связи с точки зрения отдельного организма. Он придумал понятие “совокупная приспособленность”. Я неофициально (может быть, слишком неофициально, но сам Гамильтон дал на то свое благословение) переопределил совокупную приспособленность как “показатель, который особь с виду стремится максимизировать, когда на самом деле максимизируется выживание генов”. Таблица ниже резюмирует представления о “репликаторах” и “носителях” и объясняет, как и те, и другие являются единицами естественного отбора, но в разном смысле.
В книге “Расширенный фенотип” я прибег к аналогии с кубом Неккера (см. ниже), чтобы подкрепить мысль, что оба подхода к рассмотрению естественного отбора сводятся к одному и тому же, как и оба представления куба Неккера равно согласуются с информацией, поступающей от глаз. К кубу Неккера я еще вернусь в одном из дальнейших разделов этой главы.
В “Эгоистичном гене” я заявлял, что следую Гамильтону, но он сам переключался между гено- и индивидоцентрической (то есть совокупно-приспособленческой) реализациями своей модели. Вот как он формулировал геноцентрические представления:
Естественный отбор благоприятствует гену, если совокупность его копий образует растущую долю общего генофонда. Мы будем рассматривать гены, предположительно воздействующие на социальное поведение носителей, так что попробуем оживить эти доводы, временно приписав генам разум и некоторую свободу выбора. Представим, что ген размышляет над задачей увеличения числа своих копий…
Эти слова он написал лет через восемь после статьи о совокупной приспособленности, но совершенно очевидно, что тот же геноцентрический взгляд лежал в основе его великого труда 1964 года. В “Эгоистичном гене” я тоже довольно свободно переключался с самого гена как метафорического субъекта, принимающего решения, на неформальный взгляд с точки зрения совокупной приспособленности, где я позволял отдельной особи вести монолог о том, что лучше для ее генов. Не нужно объяснять, что ни один из этих монологов не следует воспринимать буквально. Следует считать, что “субъекты” в обоих случаях ведут себя,
Гамильтон основывал свои представления на точке зрения гена, но совокупная приспособленность – это и способ сохранить привычный фокус на отдельном организме, носителе. Все эти старания вернуть индивид вместо гена в фокус нашего дарвиновского внимания видятся мне прискорбно неуклюжими. Но почему вообще отдельный организм – такой ярко выраженный носитель? Почему мы принимаем организм как должное? Мы ждем геноцентрического ответа на вопросы, почему существуют крылья и глаза, рога и пенисы, – разве не должен геноцентрический взгляд вызывать у нас и вопрос, почему существуют сами организмы? Гены выживают, дергая фенотипические рычаги управления. Но почему эти фенотипические рычаги увязаны в отдельные носители, которые мы зовем организмами, и почему бессмертные репликаторы “решаются” сколотить банду с другими генами и ехать на общем носителе? Здесь я зашел дальше Гамильтона, но при этом никогда всерьез не противоречил ничему из того, что он говорил. Эти представления я изложил в своей второй книге, “Расширенный фенотип”. У меня взял интервью Марек Кон для собственной книги, “Причина всего”, с подзаголовком “Естественный отбор и английское воображение”, где удачно описал мои доводы.
Его первая книга основывалась на допущении: “если адаптации и существуют «для блага» чего-то, то этим чем-то является ген”. Теперь Докинз готовит попытку “освободить эгоистичный ген из его концептуальной тюрьмы – индивидуального организма”.
Одним из тюремщиков в этой Бастилии был Билл Гамильтон: его записали на роль революционера, которого его ученик счел недостаточно радикальным. Докинз непоколебимо преклонялся перед Гамильтоном, но считал, что представление о совокупной приспособленности препятствует взгляду на биологические факты с точки зрения гена. Совокупная приспособленность рассматривает отбор генов, но усложняет вопрос, пытаясь вписаться в устоявшиеся рамки биологии. “До революции, которую произвел Гамильтон, наш мир был населен индивидуальными организмами, простодушно старавшимися остаться в живых и принести потомство. Успех в таком деле было логично измерять на уровне индивидуума. Гамильтон изменил все это, но, к сожалению, вместо того чтобы довести свои идеи до логического завершения и низвергнуть индивидуальный организм с пьедестала <…>, он направил свой гений на разработку средства для его спасения”[129].
Джон Мэйнард Смит сказал практически то же самое в интервью, которое я взял у него в 1997 году для “Сети историй” (см. стр. 274) [130].
Расширяя фенотип
Если святой Петр у райских врат потребует ответа, чем я могу оправдать место, которое занимал на этой земле, и воздух, которым дышал, – я не найду ничего лучше, чем указать на “Расширенный фенотип”. Это не какая-то принципиально новая гипотеза, которая может оказаться верной или ошибочной, которую нужно проверять экспериментами или наблюдениями. Скорее это новый способ смотреть на хорошо знакомое: способ смотреть на биологию так, что все раскладывается по полочкам и обретает смысл. Видимо, это нечто вроде сентенции “Сегодня первый день твоей оставшейся жизни”: банально, безусловно истинно, не то утверждение, для которого стоит отправляться на поиски доказательств, – но мы, тем не менее, признаем, что это истина, меняющая наш взгляд на вещи. Как бы она ни была очевидна, иногда стоит удосужиться произнести ее вслух, чтобы повлиять на чьи-то действия. Так я вижу расширенный фенотип. Но вместо того, чтобы сводить его к хлесткому афоризму, я хочу его объяснить. Одно из возможных объяснений – в том, что он ставит под вопрос предположительную центральную роль “носителя”.
Было принято считать, что фенотипическое влияние гена ограничено телом особи, содержащей этот ген. Гены влияют на тела посредством процессов эмбрионального развития. Так, мутантная версия гена слегка меняет форму крыла стрижа. Из-за этого птица летает чуть быстрее при тех же затратах энергии, а от этого с чуть большей вероятностью выживает и, следовательно, передает тот же самый ген будущим поколениям. Умножьте эффект на множество стрижей и множество поколений, и в результате мутантный ген будет доминировать в популяции, вытеснив альтернативные аллели[131].
Чтобы разобраться в том, каково непосредственное воздействие конкретных генов, нужно погрузиться очень глубоко в биохимию отдельного организма: на это способны лишь узкие специалисты-ученые. А вот фенотипические воздействия обычно проявлены внешне и видны невооруженным взглядом – как, например, крыло стрижа: их мы распознаем как приспособления для выживания. Но существует целый каскад скрытых, внутренних причин и следствий, который часто начинается с синтеза белка, в точности закодированного последовательностью ДНК. Мы могли бы произвольно определить “фенотип” в любой точке этого каскада: им можно назвать сам белок, или его непосредственный катализирующий эффект для клеточной биохимии, или влияние этого эффекта на поведение клеток, взаимодействующих в тканях, или множество дальнейших последствий – даже не доходя до чего-то наблюдаемого во внешнем строении животного: крупнее перепонки на лапах утки, больше крылья у осы или более неуклюжий ухаживающий жест альбатроса. Под фенотипическими воздействиями гена было бы правильно понимать это все.
Новой в “Расширенном фенотипе” была мысль, что эта последовательность причин и следствий не обязана останавливаться на границе тела. Рассмотрим, скажем, трубочки из грязи, какие строят осы Джейн Брокманн,
Да, именно так –
Откуда я знаю, что эти гены существуют? Я этого не знаю. То есть никто еще не проводил генетического исследования фенотипических черт из списка выше. Но я уверен, что если провести это генетическое исследование – а это вполне возможно, – то обнаружится, что все эти фенотипические черты меняются под управлением генов. Откуда такая уверенность? Дело в том, что трубочки, которые строит оса, очевидно, приобрели свою удачную форму под действием естественного отбора, а логика естественного отбора подразумевает участие генов. Как мог бы естественный отбор сформировать трубочки из грязи, которые все лучше выполняют свою функцию защиты личинок, если он не благоприятствовал определенным генам за счет остальных? Повторюсь, безусловно, гены влияют на трубочки лишь опосредованным образом, через строительное поведение ос. А на него (предыдущий шаг в причинно-следственной цепочке) – через нервную систему ос. А на нее – через клеточные процессы, строящие нервную систему ос. Но в любом случае все фенотипические эффекты опосредованы. Влияние генов на грязевые трубочки опосредовано точно так же, как влияние генов на крылья, лапки и усики. А расширенный фенотип, который мы рассматриваем, может оказаться и не последним звеном в цепочке причин и следствий. Все его дальнейшие последствия тоже можно считать расширенным фенотипом при единственном условии: что гены, отвечающие за него, были отобраны в ходе естественного отбора.
В этой книге приводится иллюстрация – разнообразие цвета грязевых трубочек. Значит ли это, что существуют гены, определяющие цвет трубочек? Возможно. Здесь я менее уверен, но лишь потому, что неочевидно, что цвету трубочек благоприятствовал естественный отбор. Возможно, что одни цвета лучше, чем другие, и возможно, что гены заставляют ос придирчиво выбирать грязь по цвету. С другой стороны, вполне возможно, что цвет грязи ос не волнует: они собирают любой доступный в ближайшем ручье материал, который может оказаться светло-коричневым, темно-коричневым или рыжевато-коричневым. Почему нельзя сказать, что ос также не волнует длина трубочки или толщина стенки? Может быть и так, но, кажется, это маловероятно. Легко заметить, что стенка может оказаться слишком тонкой для своей задачи (а значит, плохо защитит личинку или вовсе развалится). Или слишком толстой (а значит, потребуется больше походов к ручью за грязью, на которые тратится ценное время). Трудно вообразить, что толщина стенок трубочки могла бы
Это гипотетические черты, о которых я говорю лишь для примера. Суть в том, что логика естественного отбора (выбор генов за их фенотипические эффекты) вынуждает нас признать, что подобные функциональные фенотипы не ограничены отдельным телом, “носителем”. И самые простые и очевидные примеры нам предоставляют артефакты животных. Здесь я воспользовался близкой дружбой с Майклом Ханселлом: в бытность студентами Оксфорда мы вместе снимали квартиру. Сегодня Майк – мировой эксперт по артефактам животных, автор нескольких трудов о них, в том числе прекрасной книги “Построено животными”: в ней тема артефактов находчиво задействуется как отправная точка, чтобы поговорить в более общем смысле о разных аспектах поведения животных. В книге “Расширенный фенотип” целая глава посвящена артефактам животных: домикам личинок ручейников, птичьим гнездам, термитникам, плотинам бобров. Даже озеро, возникающее у бобровой запруды, можно с полным правом считать (расширенным) фенотипическим проявлением генов бобра – вероятно, это самый крупный в мире фенотип.
Если бы “Расширенный фенотип” ограничивался артефактами наподобие грязевых трубочек у ос Джейн Брокманн или передвижных домиков у ручейников Майка Ханселла, я бы не потрудился говорить (а издатели не стали бы печатать на задней обложке книги): “Неважно, читали ли вы у меня что-то еще, прочитайте, пожалуйста, хотя бы