Книги
Эмоциональный мозг
Демонстративным примером сложного взаимодействия симпатических и парасимпатических эффектов при эмоционально положительном состоянии может служить динамика сексуального возбуждения у человека. Хотя в картине начальной стадии возбуждения наблюдаются симпатические сдвиги (подъем кровяного давления), на этой стадии преобладает активация парасимпатической системы. После интермиссии симпатические эффекты тахикардии и гипервентиляции приобретают доминирующий характер. «Симпатический пик» сменяется фазой парасимпатической гиперкомпенсации. Признаки одновременного нарастания активности двух отделов вегетативной нервной системы встречаются гораздо чаще, чем реципрокные отношения, а моменты относительного преобладания одного из отделов отнюдь не означают полного доминирования. Хотя динамика вегетативных сдвигов в определенной мере зависит от двигательной активности, эти сдвиги не могут быть сведены к вегетативному «аккомпанементу» моторики. Высокая степень эмоционального возбуждения одного из сексуальных партнеров может вызвать сильнейший сдвиг вегетатики (учащение сердцебиений до 170 уд./мин) у второго партнера в отсутствии движений. Таким образом, динамика вегетативных сдвигов при сексуальном возбуждении имеет сложную внутреннюю структуру, непосредственно не коррелирующую с двигательной активностью субъекта.
И веселье, и грусть сопровождаются у человека активацией симпатического отдела, причем для грусти более характерны сдвиги сердечно-сосудистой системы, а для веселья — изменения дыхания. Симптомы возбуждения симпатического отдела в виде учащения пульса, повышения кровяного давления и температуры, уменьшения слюноотделения и электрического сопротивления кожи характерны для многих отрицательных эмоций. Вместе с тем в структуре этих реакций обнаруживаются признаки активации парасимпатического отдела. Степень участия симпатических и парасимпатических влияний зависит от характера данной отрицательной эмоции. Активно-оборонительные агрессивные реакции обезьян сопровождаются учащением сердцебиений, пассивно оборонительные — брадикардией [Джелиев, Лагутина, Фуфачева, 1963]. Аналогично влияние эмоций и на сердце человека: агрессивные, равно как и положительные по своей окраске реакции реализуются на фоне учащения пульса, состояния пассивности и депрессии имеют тенденцию к урежению сердцебиений [Theorell, Blunk, Wolf, 1974].
Итак, анализ литературных данных приводит нас к выводу о том, что симпатический и парасимпатический отделы вегетативной нервной системы вовлекаются в реализацию как отрицательных, так и положительных эмоциональных состояний. Любая эмоциональная реакция характеризуется своим типом взаимодействия симпатических и парасимпатических влияний. В каждом конкретном случае возникновения эмоционального напряжения складывается именно та комбинация симпатических и парасимпатических влияний, которая оказалась наиболее целесообразной и была закреплена естественным отбором. Так называемая «саморегуляция» сердечно-сосудистой системы имеет лишь подсобное значение ограничителя слишком больших отклонений в процессе эмоциональной мобилизации вегетативно-энергетических ресурсов организма.
На рис. 14 мы попытались схематически изобразить динамику активации симпатического и парасимпатического отделов вегетативной нервной системы по мере роста положительного и отрицательного эмоционального напряжения. В основу схемы положена классификация базальных эмоциональных состояний, о которой мы будем говорить в последней главе. Нам было важно подчеркнуть, что в процессе реализации эмоций можно встретить и синергизм, и реципрокность симпатических и парасимпатических влияний на вегетативные функции организма. Мы сознаем, что схема груба и требует дальнейших уточнений.
Изменения электроэнцефалограммы при эмоциональных реакциях человека
Степень эмоционального напряжения нередко рассматривают как феномен, идентичный уровню бодрствования. Согласно этим представлениям эмоциональное возбуждение занимает в континууме бодрствования крайнее положение, диаметрально противоположное глубокому медленноволновому сну и коме. Подобный взгляд на природу эмоционального возбуждения, подкрепленный открытием функций ретикулярной формации мозга, получил свое наиболее законченное воплощение в «активационной теории эмоций» Дональда Б. Линдсли [1960]. Однако факты демонстрируют многочисленные примеры явной диссоциации между степенью эмоционального возбуждения, если о нем судить по сдвигам вегетативных функций, и уровнем бодрствования, диагностированным по суммарной электрической активности мозга. Ожидание болевых раздражений в опытах Р. Фроста с соавторами приводило к сдвигам частоты сердцебиений и электросопротивления кожи без достоверных изменений альфа-ритма электроэнцефалограммы — ЭЭГ [Frost, Burish, Holmes, 1978]. В состоянии страха, при восприятии эмоционально окрашенных слов, во время сдачи экзаменов были зарегистрированы не угнетение, а усиление альфа-ритма, повышение его амплитуды, возрастание альфа-индекса.
Вопрос о соотношении вегетативных и ЭЭГ признаков эмоционального возбуждения был подвергнут систематическому анализу в экспериментах М. Н. Русаловой [1979]. На рис. 15 хорошо видно, что эмоциональное напряжение, о котором судили по учащению сердцебиений, может сопровождаться у одного и того же субъекта как угнетением, так и экзальтацией альфа-ритма. Чем различаются эти две ситуации? В первом случае субъект ожидал реальное болевое раздражение кожи предплечья. Во втором — ему была дана инструкция мысленно представить болевое раздражение в конце падающего отсчета, записанного на магнитофон.
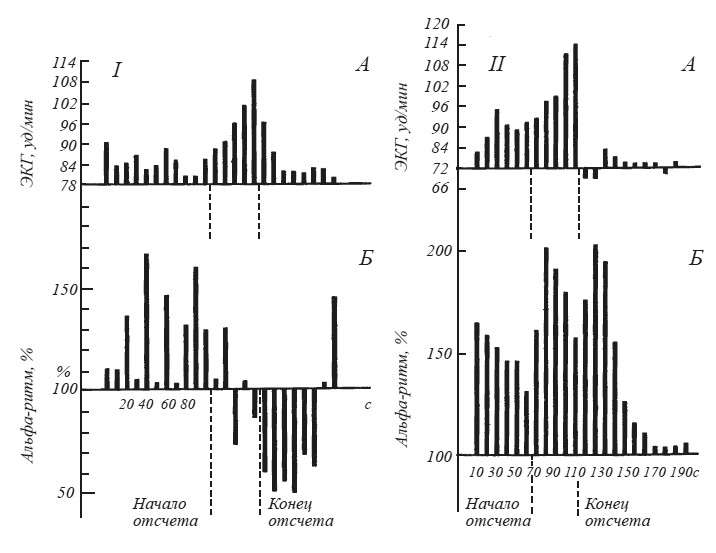
Рис. 15. Динамика изменений амплитуды (Б) альфа-ритма (по показанию пера интегратора) и частоты сердечного ритма (Д) после инструкции, в период отсчета и после отсчета
Альфа-ритм (11 Гц) выражен в процентах к фону, принятому за 100;
Выше мы говорили, что эмоциональная реакция может возникнуть как при поступлении информации из внешней среды, так и при извлечении следов ранее полученных впечатлений из памяти. Многочисленные эксперименты, поставленные М. Н. Русаловой (Валуевой), равно как и данные, почерпнутые в литературе, убеждают в том, что депрессия альфа-ритма характерна для ситуаций, в которых внимание субъекта обращено к окружающей его внешней среде. Если субъект преимущественно сосредоточен на следах, хранящихся в памяти, даже высокая степень эмоционального возбуждения не ведет к угнетению альфа-ритма, а зачастую сопровождается его экзальтацией. Подобный вывод хорошо согласуется с представлением об альфа-ритме как электрофизиологическом корреляте механизма, квантующего поток поступающей в мозг афферентации [Walter, 1954]. Нуну и Осселтон показали, что короткие зрительные стимулы не воспринимаются, если они подаются во время определенных фаз альфа-ритма. По мнению авторов, альфа-ритм коррелирует с деятельностью «нервной задвижки», которая периодически прерывает поступление информации в мозг [Nunn, Osselton, 1974].
Может возникнуть вопрос: почему же при сильном потоке афферентации, когда необходимость в работе «задвижки» особенно велика, альфа-ритм исчезает, депрессируется? Здесь не следует отождествлять деятельность механизма квантования с выраженностью его электрофизиологических коррелятов. Хорошо регистрируемый альфа-ритм при закрытых глазах, тишине и покое — это «холостой ход» квантующего механизма, который, кстати, существует и у лиц без выраженного альфа-ритма в ЭЭГ. Сказанное справедливо и для динамики тета-ритма, о котором сейчас пойдет речь.
Одним из ЭЭГ, симптомов эмоционального возбуждения служит усиление тета-ритма с частотой колебаний 4–7 Гц. С помощью анализаторов частотного спектра ЭЭГ тета-ритм был зарегистрирован у здоровых людей при отрицательных [Суворова, 1975] и положительных [Walter, 1953; Валуева, 1967] эмоциях. У детей, занятых деятельностью, окрашенной положительными эмоциями, тета-ритм лучше выражен в правом полушарии большого мозга, особенно в лобных его областях [Денисова, 1978]. Для понимания функционального значения тета-ритма интересны данные Брауна [Brown, 1971], который демонстрировал субъекту альфа-, бета- и тета-полосы его собственной ЭЭГ в виде различной цветовой окраски экрана. Испытуемых спрашивали, какому их субъективному состоянию соответствует тот или иной цвет. Оказалось, что для тета-ритма характерны обдумывание планов, неопределенность мысленно анализируемой ситуации, изменение темы размышлений, решение технических и житейских проблем, «сны наяву». Нарастание процентного содержания тета-ритма в спектре ЭЭГ совпадает со снижением бдительности человека-оператора к внешним сигналам [Beatty, Greenberg, Deibler, O’Hanlon, 1974].
Для того чтобы оценить функциональное значение изменений ЭЭГ при эмоциях, остановимся на гипотетическом механизме ритмических колебаний биопотенциалов мозга. Согласно гипотезе П. Андерсена и Дж. Экклса, важную роль в формировании этих колебаний играют тормозные нейроны [Andersen, Eccles, 1962]. Это дает основание применить к взаимодействию возбужденных и тормозящих их деятельность нервных микроструктур теорему В. Вольтерра [1976] о закономерных изменениях амплитуды и частоты осцилляций в системе, состоящей из двух типов функционально антагонистических элементов (табл. 3).
Таблица 3. Изменение электроэнцефалограммы, предсказываемое теоремой Вольтерра
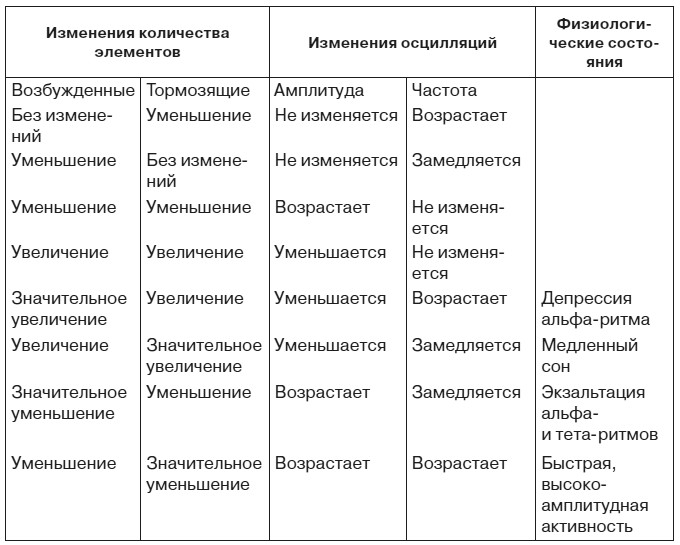
В какой мере картина, представленная в таблице, совпадает с физиологическими фактами? Наиболее бесспорен, пожалуй, механизм классической депрессии альфа-ритма, которая определяется нарастанием числа возбужденных и тормозящих элементов с преимущественным увеличением первых из них. Весьма неожиданным может показаться тот факт, что картина, характерная для медленного сна, также обусловлена увеличением возбужденных и тормозящих элементов, только число последних нарастает в большей мере, чем количество первых. Впрочем, данные последних лет не позволяют отвергнуть этот вывод. Усиление активности нейронов коры и ретикулярной формации во время сна побуждает рассматривать сон скорее как результат перестройки взаимодействия нервных элементов, чем как состояние «разлитого торможения» коры и подкорки в старом понимании этого термина. Ряд авторов придают важное значение нарастанию активности механизмов возвратного торможения в генезе медленного сна.
Высокоамплитудные медленные колебания свидетельствуют об уменьшении количества и возбужденных, и тормозящих элементов с относительным преобладанием последних. Уменьшение количества возбужденных элементов, но с преимущественным уменьшением тормозящих микроструктур имеет место и при быстрой высокоамплитудной активности. Применение теоремы Вольтерра к ритмической биоэлектрической активности мозговых структур не только создает предпосылки для математического моделирования ЭЭГ, но и побуждает по-новому взглянуть на внутренний нейрональный механизм ряда типичных изменений электроэнцефалограммы.
Если в генезе альфа-ритма решающее значение имеют кортикоталамические отношения, то тета-ритм связан с кортико-лимбическим взаимодействием. Появление тета-ритма в ЭЭГ человека, находящегося в коматозном состоянии, можно рассматривать как результат активности филогенетически древних лимбических структур на фоне деактивации менее устойчивых неокортикальных образований [Гриндель, Брагина, Добронравова, Доброхотова, 1974].
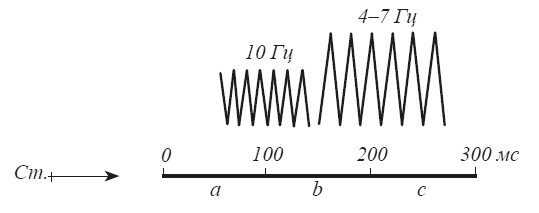
Итак, альфа-ритм — это «сенсорный» ритм, связанный с поступлением информации из внешнего мира. Не случайно он лучше всего выражен в зрительных отделах новой коры, поскольку у человека зрение является основным каналом получения информации об окружающей внешней среде. «Лимбический» по своему происхождению «эмоциональный» тета-ритм, по-видимому, имеет какое-то отношение к деятельности «оценивающих» механизмов мозга. С таким предположением хорошо согласуются временные параметры альфа- и тета-ритмов. Согласно данным психофизики восприятия первые 100 мс необходимы для отражения в структурах мозга физических характеристик стимула, для формирования его «нервной модели». На протяжении следующих 100 мс мозг производит сопоставление модели с энграммами, извлеченными из памяти. Между 200–300 мс выясняется результат сличения и начинается подготовка к реакции, если наличный стимул требует ответного действия [Ломов, Иваницкий, 1977]. Продолжительность первого этапа совпадает с временными характеристиками альфа-ритма. Продолжительность процессов сличения и «принятия решения» (150–250 мс) идентична частоте тета-ритма (рис. 16).