Книги
С нами или без нас: Естественная история будущего
Макартур помог Уилсону проработать и расширить эти идеи, а также запечатлеть их в наборе уравнений. Результатом их совместного труда стала книга «Теория островной биогеографии» (The theory of island biogeography). Впоследствии их теорию тестировали на самых разных островах, разбросанных по морям и океанам. Ее проверяли десятки, а потом и сотни ученых, в основном студенты и аспиранты, стремящиеся разобраться в скрытых правилах, по которым живет наш мир. Детали сформулированных Макартуром и Уилсоном уравнений изучались и оспаривались с дотошностью, какую ученые обычно приберегают для самых важных поводов. По ходу дела было выяснено, что уравнения не учитывают многие свойства биологии островов; тем не менее теория выдержала проверку временем. Она отражает важнейшие нюансы того, как устроен наш мир: на крупных островах действительно живет больше видов, и это действительно объясняется соотношением вымираний и возникновений. Возможно, не менее важно и еще одно обстоятельство: теория Макартура и Уилсона содержит четкие прогнозы относительно того, каким окажется будущее природы, – идет ли речь об отдаленных островах, диких лесах или даже городах. Особенно о городах.
Экологам потребовалось не слишком много времени, чтобы понять: теория Макартура и Уилсона может применяться и к тем изолированным средам обитания, которые похожи на острова, – а таковых в наши дни можно найти довольно много. В конце концов, так ли уж отличается клочок британского леса, со всех сторон окруженный морем сельскохозяйственных угодий, от какого-нибудь скалистого клочка суши в настоящем море?{22} И не образуют ли зеленые разделительные полосы посреди Бродвея на Манхэттене нечто вроде архипелага в океане стекла и бетона? Более того, распространение идей Макартура и Уилсона на изолированные среды обитания оказалось более чем своевременным. Исчезновение лесов и прочих диких ландшафтов шло угрожающими темпами. И если теория этих ученых относительно островов оказывалась применимой к исчезающим лесам, то ее положения можно было распространить и на виды, там обитающие. Но возможно ли в разрозненных фрагментах рассмотреть целостный нарратив? Макартур и Уилсон полагали, что да. Из этой убежденности выросло много масштабных исследовательских проектов, в том числе беспрецедентный эксперимент по целенаправленному воспроизводству лесных участков в бразильской Амазонии, которым руководил Том Лавджой, сотрудник Смитсоновского института.
Терри Уильямс, размышляя о нашей планете, писала: «Если мир развалится на куски, мне нужно будет понять, можно ли из его фрагментации извлечь какую-нибудь историю»{23}. К тому же стремился и Лавджой: ему важно было разобраться, какие последствия может иметь раздробленность сред обитания. В ходе возглавляемого им эксперимента фрагменты леса обособлялись путем преобразования окружающих их участков земли в пастбища. Этот лес в любом случае предназначался к вырубке, лесники извели бы его по одному деревцу, и поэтому Лавджой смог убедить бразильские власти превратить эту вырубку в научный опыт. Датский глагол «разрезать» (
Хотя детали и динамика того, каким образом утрата сред обитания влияет на биоразнообразие, по-прежнему уточняются исследователями, мы уже знаем достаточно, чтобы начать действовать{25}. Уилсон и другие биологи-консервационисты призывают сохранить половину земной суши в виде диких лесов, лугов и других экосистем. По мнению Уилсона, именно столько нужно для того, чтобы сберечь биоразнообразие, в котором мы нуждаемся сейчас или будем нуждаться в будущем. И он знал, о чем говорил, ведь с его помощью было сформулировано заветное уравнение, упоминавшееся выше.
Динамику островной биогеографии почти всегда можно спрогнозировать довольно точно: для этого достаточно принимать в расчет появление или исчезновение на острове или в ином изолированном фрагменте новых видов – то есть колонизацию и вымирание. Однако здесь играет роль и еще один процесс, ранее упомянутый Макартуром и Уилсоном, но в дальнейших исследованиях затрагивавшийся редко. Речь идет о видообразовании.
Видообразование – это появление новых видов, возникновение двух или более видов там, где раньше был только один. Считается, что скорость видообразования возрастает вместе с увеличением площади среды обитания. Изначально Макартур и Уилсон предполагали, что не только на крупные острова прибывает больше новых видов, но и видообразование там более вероятно и происходит быстрее. В годы, последовавшие за публикацией в 1967 г. «Теории островной биогеографии», это предсказание почти не обсуждалось. Возможно, гипотезы Макартура и Уилсона о видообразовании остались без внимания из-за того, что они были изложены на самых последних страницах книги. Но не исключено, что они просто опередили свое время. Экологи и эволюционные биологи на тот момент еще не осознавали, насколько быстро способна идти эволюция, и тем более не представляли, что возникновение видов можно задокументировать в реальном времени.
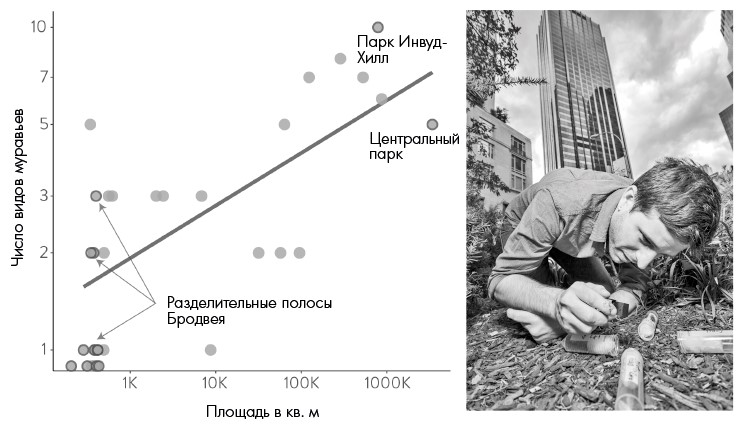
Рис. 2.1. Слева: пример соотношения разнообразия видов и площади сред обитания, аналогичных островам, – муравьи на разделительных полосах и в парках Манхэттена. Справа: Клинт Пеник (на тот момент научный сотрудник в моей лаборатории, а теперь старший преподаватель Государственного университета Кеннесо) собирает образцы муравьев на одной из разделительных полос, заманивая их сахаром в небольшие колбы. Диаграмма составлена Лорен Николс по данным из: Savage, Amy M., Britné Hackett, Benoit Guénard, Elsa K. Youngsteadt, and Robert R. Dunn, "Fine-Scale Heterogeneity Across Manhattan's Urban Habitat Mosaic Is Associated with Variation in Ant Composition and Richness," Insect Conservation and Diversity 8, no. 3 (2015): 216–228.
Тот, кто дочитает книгу Макартура и Уилсона до конца, обнаружит, что авторы обсуждают тему видообразования довольно подробно. Они подчеркивают, что острова – «прекрасное поле для изучения эволюции»{26} во всем, что касается видообразования, адаптации к местности или просто возникновения новых черт. Взгляд на острова как на эволюционную арену связывал Макартура и Уилсона с Дарвином. Для Дарвина острова одновременно были призмой, сквозь которую он исследовал эволюцию, и пространством, на котором он проверял свои размышления. Изолированные участки суши, которые Дарвин посетил за почти пятилетнее путешествие на корабле «Бигль», – в том числе Кабо-Верде, Фолкленды, Галапагосы, Таити, Новая Зеландия и континент-остров Австралия – позволили ему обстоятельно изучить целый круг видов, которые он не встречал нигде больше. Как он осознал позже, во многих случаях заинтересовавшие его виды появились и эволюционировали именно на этих островах. Острова также предлагали идеальный контекст, в котором удобно было описывать ход естественного отбора: разграничить его на фазы, отталкиваясь от которых можно видеть процесс, идущий повсеместно.
Как писал Дарвин, новые виды формируются на островах в ответ на изоляцию и специфические местные условия. Острова Галапагосского архипелага – это древние вулканы, поднявшиеся с океанского дна в 800 километрах от западного побережья Южной Америки. Когда-то сюда прибыл единственный вид черепахи средних размеров – и со временем он эволюционировал в целых 14 видов гигантских черепах: одни побольше, другие поменьше, одни потемнее, другие посветлее. Один-единственный вид пересмешников прилетел на архипелаг – и эволюционировал в три вида, каждый из которых обитает на отдельном острове. Один легко мутирующий вид серых вьюрков тоже достиг островов – и эволюционировал в 13 видов, их теперь называют дарвиновскими вьюрками. Дарвин заметил, что у здешних вьюрков различались клювы. В «Путешествии натуралиста вокруг света на корабле "Бигль"» он писал, что посредством естественного отбора «на этом архипелаге был взят один вид и видоизменен в различных целях»{27}. В частности, один из видов галапагосских вьюрков эволюционировал так, что клювы его представителей могли доставать нектар, пыльцу и семена из кактусов. Другой вид превратился в вампиров, клюющих в спину птиц и других позвоночных, чтобы пить их кровь. Еще у двух видов появилась способность удерживать в клювах палочки, с помощью которых они охотились на личинок. Наконец, у некоторых видов развились клювы, подходящие для поедания семян.
Дарвин предполагал, что на океанских островах должны с большей вероятностью обнаруживаться эндемичные виды – то есть такие, которые не встречаются нигде больше. Ученый понял, что подобные виды возникли благодаря изоляции, из-за которой у них развились особенности, отличающие их от материковых родственников. Но у Дарвина не сложилось однозначного представления, на каких островах новые виды появляются в большем, а на каких в меньшем количестве. Макартур и Уилсон дополнили классический дарвиновский сюжет островной эволюции. Их вкладом стала гипотеза о том, что организмы, попавшие на острова, дают тем больше видов, чем крупнее остров. Однако проверить это предположение оказалось непросто. Собственно говоря, по состоянию на 2006 год оно оставалось почти непроверенным – если не считать одного графика, напечатанного в книге Макартура и Уилсона. На этой картинке, «диаграмма 60», авторы приводят количество видов птиц на островах разной площади, встречающихся только там. На диаграмме не слишком много точек, но те, что есть, действительно заставляют прийти к выводу, что на более крупных островах больше эндемических видов птиц – возможно, из-за того, что там они и сформировались.
В 2006 году Яэль Ки́сел поступила в аспирантуру Имперского колледжа Лондона. Она работала с Тимом Барраклоу, который теперь занимает профессорскую должность в Оксфорде. Кисел предстояло провести самое амбициозное и обширное исследование, посвященное тому, как площадь острова сказывается на эволюционировании на нем новых видов. На протяжении миллионов лет из океана поднимались вулканические острова. Лава кипела, а затем остывала. Там селились водоросли. Там селились птицы. Пауки выпускали шелковые нити и уносились ветрами; приземлившись на новую сушу, тоже селились там. Растения путешествовали на птичьих лапках и по водным потокам. А затем в дело вступала эволюция, условия которой задавались как местными обстоятельствами, так и прибывшими видами. Именно ее итоги и должна была изучить Кисел.
Ее работа начиналась как побочный проект. Когда она занималась своей диссертацией, Барраклоу предложил ей попутно выяснить, до какой степени маленьким может быть остров, чтобы из одного вида произрастающих на нем растений могли бы со временем появиться два. Это начинание предлагалось базировать на аналогичном исследовании, незадолго до того проведенном на птицах{28}. Как пояснила мне Кисел в электронной переписке, она пыталась понять, существует ли «какой-то минимум площади, который делает остров пригодным для видообразования у растений», и если да, то каков он. По ходу дела Кисел и Барраклоу решили расширить проект и включить в него другие виды организмов. В итоге Кисел продолжала собирать информацию до тех пор, пока не обнаружила, что ей удалось сформировать самую крупную на тот момент базу данных, описывающую свойства островов, на которых шло видообразование разных групп организмов. Причем все сведения она собрала, не покидая пределов Европы; ей не пришлось посещать ни Галапагосы, ни Реюньон, ни Мадагаскар. Оказалось, что требуемую работу можно было проделать, опираясь исключительно на музейные собрания и компьютерные базы данных, составленные теми, кто выезжал «в поле».
База Кисел содержала данные не только о небольших океанских островах, например Галапагосских, но и об островах покрупнее: самым солидным был Мадагаскар. Предметом ее исследований могли стать два типа видообразования. Прежде всего можно было бы сфокусироваться на том, будет ли некий прибывший на остров вид эволюционировать в новый, отличающийся от его материковых родственников. Но Кисел и Барраклоу интересовало в первую очередь кое-что другое – видообразование, которое разворачивается в пределах островов. Сосредоточившись именно на этом, Кисел могла не только вычислить минимальный размер острова, необходимый для видообразования (то есть разрешить исходную проблему), но и вскрыть другие потенциально значимые для видообразования факторы.
В процессе работы обнаружилось, что размер острова действительно влияет на вероятность появления новых видов, как и ожидалось по теории Макартура и Уилсона. В каждой группе организмов, рассмотренной исследовательницей, он выступал единственным значимым фактором, определяющим вероятность видообразования. Чем больше был остров, тем большей оказывалась вероятность того, что на нем образуются виды. Но этим дело не ограничилось. Основываясь на предшествующих исследованиях и собственных наблюдениях, Кисел сформулировала гипотезу, согласно которой организмы, не слишком приспособленные к перемещению между островами или по ним, будут более склонны к видообразованию на маленьких островах. И наоборот, организмы, которые с легкостью расселяются (и тем самым распространяют свои гены повсюду), редко или никогда не произведут на подобных островах новые виды.
Логика, которой руководствовалась Кисел, была вполне обоснованной. Организмы, которые беспрепятственно распространяются – быстро летают, далеко бегают или хотя бы прытко ползают, могут на какое-то время изолироваться в разных частях условного небольшого острова. Со временем, однако, существа с одной части острова неизбежно встретятся с существами с другой части. Они будут скрещиваться и обмениваться генами, а это сгладит все различия, которые успели сформироваться у популяций. Давайте представим гипотетическую ситуацию: мы решили расселить на каком-то острове одичавших собак двух разных пород – причем одну породу, скажем бульдогов, оставили на одном его краю, где среда сурова и требует адаптации, а собак другой породы, например золотистых ретриверов, выпустили в более щадящую среду на другом краю. Если остров мал, а естественных препятствий на нем немного, то некоторые ретриверы неизбежно доберутся до бульдожьей части острова, а бульдоги обязательно навестят владения ретриверов; породы скрестятся и дадут потомков, имеющих гены обоих родителей. Как писал о таких ситуациях Дарвин, «всякая наклонность к модификации подавляется скрещиванием с неизмененными иммигрантами, часто прибывающими из своей родной страны»{29}. Но зато если остров достаточно велик, то две популяции собак, возможно, никогда и не встретятся. Со временем они эволюционируют, каждая своим путем, и уже не смогут скрещиваться, так что даже если вдруг они найдут друг друга, то существовать им все равно придется врозь. Коротко говоря, Кисел предсказывала, что не слишком мобильным организмам для видообразования хватит и небольших островов. Вместе с тем для хороших летунов, например летучих мышей, или хороших бегунов, например млекопитающих отряда
Барраклоу и его напарница рассмотрели эту расселительную гипотезу применительно к различным видам из обширной базы данных Кисел – птицам, улиткам, цветковым растениям, папоротникам, бабочкам и мотылькам, ящерицам, летучим мышам и хищным млекопитающим. Сборная солянка, но зато для каждой формы жизни имелись готовые данные. За рамками анализа осталось большинство млекопитающих и насекомых, а также все микроскопические организмы. В отношении каждой формы жизни, которую изучили Кисел и Барраклоу, выяснилось одно и то же: новые виды с большей вероятностью образуются на более крупных островах. Но минимальный размер острова, обеспечивающий видообразование, меньше для тех организмов, которые плохо перемещаются (улиток), и больше для тех организмов, которые делают это с легкостью (птиц или летучих мышей). Минимальная площадь, нужная улитке, чтобы породить новый вид, весьма мала: меньше квадратного километра – примерно как завод «Тесла» во Фремонте (Калифорния). А для летучей мыши с ее способностью к дальним перемещениям необходимая площадь много больше: несколько тысяч квадратных километров – это весь Нью-Йорк с пятью его округами.
Завершив проект, посвященный эволюции новых видов на островах, Кисел переключилась на другие темы, оставив множество непроверенных гипотез. Одна из них, в частности, касалась улиток. О них вообще редко вспоминают, но новые виды улиток тем не менее постоянно возникают на островах по всему миру. Улитки дают новые виды с примечательной легкостью – возможно, как раз из-за того, что перемещаются так медленно («Я смогу, смогу, смогу!»[4]). Но Кисел предположила, что дело не только в этом. Как она объяснила мне в электронном письме, чтобы порождать новые формы на островах, видам требуются два свойства. Во-первых, чтобы избежать скрещивания со своими родственниками с других островов или с материка, они должны быть домоседами. Во-вторых, для начала им нужно как-то попасть на острова. Улитки подходят под оба критерия. Их передвижения ограничены крайне малыми расстояниями и предельно низкими скоростями: за всю свою жизнь иная улитка не продвинется и на метр. Но время от времени – по крайней мере, достаточно часто, чтобы оказаться в островной среде, – эти создания перемещаются на большие дистанции: на птичьих лапах, в птичьих кишках или на бревнах-плавунах. В происхождении видов улиткам отведена золотая середина. А вот, скажем, лягушки, оказавшись на острове, с большой вероятностью будут образовывать новые виды, но попадают они туда лишь изредка. Как заметил Дарвин, перемещения за моря даются им с трудом. Поэтому лишь на немногих океанских островах имеются местные виды лягушек.
Встречаются виды, имеющие скромный расселительный потенциал, сумевшие при этом преодолеть значительное расстояние до далекого острова. Такое сочетание может предполагать, что изначально вид легко расселялся, но, оказавшись на острове, утратил эту способность. Потеря способности к распространению может оказаться преимуществом для вида, если при прочих равных ему выгоднее оставаться на острове, чем сменить дислокацию. Такое происходит довольно часто, и именно так произошло с летучими мышами в Новой Зеландии. Прибыв в свое время в благодатную Новую Зеландию, окруженную к тому же враждебными морями, летучие мыши утратили способность летать. Став нелетучими, они повысили свои шансы на освоение различных сред обитания в Новой Зеландии – и воспользовались этим. На многих островах подобное происходило и с птицами. Нелетучесть многократно возникала у линий островных птиц, а после ее появления птицы часто давали множество видов. Сегодня такие птицы встречаются крайне редко – отчасти потому, что с прибытием на острова людей они становились легкой добычей либо для человека, либо же для видов, которые он привозил с собой, в том числе мышей и крыс.
Выводы и прогнозы Кисел и Барраклоу позволяют переосмыслить все то, что теория островной биогеографии рассказывает об окружающей нас жизни. Следует ожидать, что на сокращающихся по всему миру участках лесов, лугов и болот будут вымирать древние виды. Так и происходит. На некоторых из этих фрагментов также будут возникать новые виды, начало которым положат популяции, оказавшиеся изолированными от своих родичей. Но возникновение новых видов будет происходить намного реже, чем исчезновение старых, – из-за того, что вымирание вообще идет быстрее видообразования, и из-за того, что видообразование менее вероятно на небольших участках среды.