Книги
Павлов И.П. Полное собрание сочинений. Том 2. Часть 1.
С целью разъяснить это я произвел новый ряд опытов. Вначале дело велось совершенно так, как это описано при основном ряде опытов. Спустя час-полтора часа раздражение прекращалось. Слюна, однако, продолжала выделяться сама собою, постепенно замедляясь, чтобы через полчаса-часполтора часа прекратиться совсем. Часа 3-4 спустя после начала опыта курарное отравление проходило настолько, что искусственное дыхание делалось уже ненужным. Животное, однако, оставалось привязанным на столе вплоть до конца опыта. Восемь часов спустя по прекращении отделения оно убивалось кровопусканием. Из всех этих животных два представили особенность. У них отделение по прекращении раздражения продолжалось с значительной энергией вплоть до конца опыта --- 8 часов после раздражения. Когда курарное отравление проходилони сильным и порывистым дыханием давали знать о возбужденном состоянии. Нужно думать, что возбужденное состояние центральной нервной системы, вызванное сильным чувствительным раздражением, у одних собак утихает довольно скоро, у других же, благодаря особенностям их нервной системы, продолжается чрезвычайно долго, за период существовавшего внешнего раздражения. Эти две собаки, понятно, не вошли в свой десяток и были заменены другими. К ним мы еще вернемся в заключительных рассуждениях.
С железами и слюной этого рода поступлено было совершенно так же, как и ранее (табл. 5).
Отделение в каждом опыте в среднем продолжалось 2 часа 20 мин., причем 1 час 24 минуты приходятся на отделение во время искусственного раздражения, 56 минут - на произвольное отделение по прекращении внешнего раздражения. Каждый опыт продолжался по прекращении всякого отделения в среднем 7 часов 20 минут. Всего десятью железами выделено слюны 701 г в среднем, следовательно каждой железой - по 70 г.
Вот готовая таблица (5) результатов этого ряда.
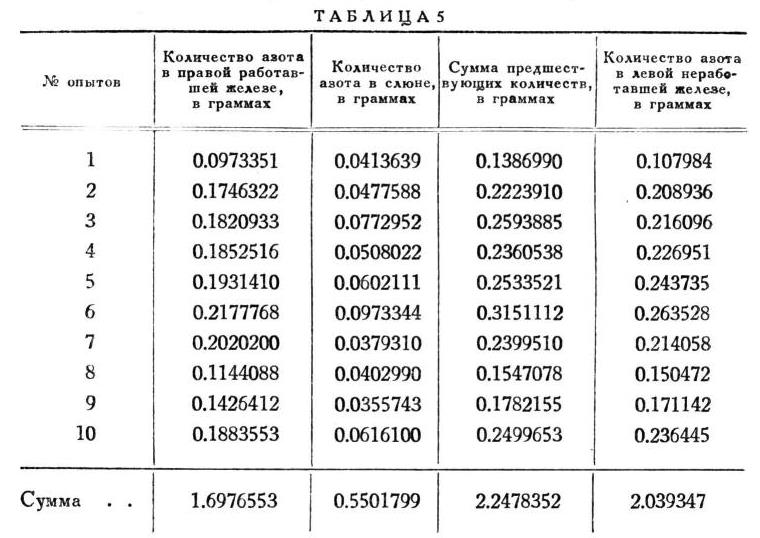
Количество азота слюны составляет 27 % количества азота неработавшей железы. Избыток азота на стороне работавшей железы равняется 0.208488 г. Это составляет 37.9% азота, потерянного правой железой при работе, и 10% всего первоначального количества его в этой железе.
Основной смысл этих цифр вполне определенный.
Мы имеем в опыте два периода: период раздражения (берем вместе как искусственное отраженное, так и автоматическое раздражение) - выделения слюны период отсутствия раздражения - отделительного покоя. Последний период втрое больше первого по продолжительности, тe. три четверти времени опыта приходится на период покоя, а периоду раздражения остается одна четверть. Мы знаем уже из прежнего ряда опытов размер восстановления железы за период полутора-двухчасового раздражения, а именно, немного более одной четверти всей потери. В настоящем ряде восстановление за все время опыта (периоды раздражения и покоя вместе) составляет немного более 1 / всей потери, т. е. больше на /10 всей потери. Будь восстановление результатом времени, оно должно бы быть в нашем последнем ряде равным потере, т. е. полным, так как время для него было теперь в 5 5 раз больше, чем в прежнем ряде, а вся продолжительность опыта в 4 раза больше продолжительности периода раздражения.
Мы должны заключить, что восстановление из двух предположенных факторов - времени и раздражения - находится в прямой зависимости только от последнего.
Но восстановление в последнем ряде, все же, было больше, чем в прежнем. Едва ли это может быть случайным колебанием при нашем массовом, коллективном опыте. И мне кажется, что возможное основание для этого излишка нетрудно заметить в условиях опыта. Только что выведено заключение, что раздражение непосредственно влияет на восстановление. Следовательно, колебания в силе или другие какие-либо особенности в своистве раздражения должны неминуемо изменять размер восстановления. А в последнем ряде, вместе с искусственным раздражением, имело место и автоматическое, наверное во многом отличающееся. от искусственного, например в том, что оно с течением времени не росло, как искусственное, а постепенно ослаблялось до нуля. Совершенно естественно, что и процент восстановления теперь оказался другим, чем в прежнем ряде.
На этом я кончаю пока мои фактические данные, но позволю себе прибавить еще некоторые теоретические рассуждения, выдвигающие ближайшие вопросы этой области.
В приведенных таблицах мы видели, что азот, потерянный железой во время раздражения отделительных нервов, возместился за то же время maximum только на 38%. Так как опыты вместе с тем показали, что полный покой железы ничего не прибавляет к величине восстановления, то поднимается естественный вопрос: когда же, при каких условиях может произойти и действительно происходит полное восстановление истраченных материалов? Для разъяснения этого вопроса мы должны обратиться к деятельности пищеварительных желез в нормальных условиях. То же отношение разрушительных и восстановительных процессов, которое мы нашли в опытах с искусственным раздражением, наблюдается и при нормальных условиях, но только в начальном периоде пищеварительного процесса. Для первых часов пищеварения постоянно всеми наблюдателями установлено преобладание разрушительного процесса над восстановительным: клеточки разных пищеварительных желез резко уменьшаются в объеме. Но затем постепенно выступают признаки восстановления клеточек, и к 24 часам или ранее после еды клеточки оказываются во всех отношениях совершенно такими, какими сни были перед едой. Какие же обстоятельства дают с течением времени перевес восстановительным процессам перед разрушительными? Как мне кажется, возможны только два предположения. Во-первых, можно иметь в виду следующее. Несколько часов спустя после начала пищеварения наступает переполнение крови и лимфы новыми питательными веществами, и это обилие свежего и свободного материала может обусловить гораздо более значительное восстановление, чем какое существует в начале пищеварения и какое наблюдали мы в наших опытах. Во-вторых, можно бы думать, что с течением пищеварительного процесса меняется степень раздражения отделительных нервов, и это изменение влечет за собою иное отношение разрушительного и восстановительного процессов. При сравнении этих двух предположений большая вероятность правды оказывается на стороне последнего. Когда мы имеем перед собою нормальные условия, т. е. когда в начале пищеварения начинает работать и, значит, разрушаться целая масса желез, тогда бы еще естественно допустить, что для полного, одновременного с разрушением восстановления их нехватает, так сказать, наличного материала в крови, и отсюда возникает преобладание разрушения железистой клеточки над ее восстановлением. Но я в моих опытах имел дело с усиленною деятельностью только четырех маленьких желез (слюнные железы другой стороны, за исключением околоушной. не отделяли вследствие перерезки барабанной струны); и едва ли можно сомневаться, что для восстановления потерь этих органов в крови, наверное, должно было оказаться достаточное количество материала, тем более, что употребленные в дело собаки бывали и в средних ступенях пищеварения, но отнюдь не голодавшие, т. е. не евшие более 20-24 часов. Когда случались собаки еще во время пищеварения, то для восстановления слюнных желез должны были существовать исключительно благоприятные условия, ибо при обилии свежего материала в крови значительная часть пищеварительных (желудочных, поджелудочных и т. д.) желез, наверное, не работала в период раздражения седалищных нервов вследствие доказанного угнетающего влияния такого раздражения на эти железы. Таким образом едва ли можно в колебаниях содержания свежего материала в крови искать объяснение тому, что в начале пищеварения в железах преобладает разрушение, а позднее - восстановление.
Второе предположение более согласуется с наличными фактами и наблюдениями. В настоящее время имеются кривые нормального отделения почти для всех пищеварительных желез. На этих кривых видно, что скорость отделения правильно изменяется в течение пищеварительного периода: к 2-3 часам. После начала пищеварения скорость отделения достигает maximum а, а затем вообще начинает постепенно падать. Если мы возьмем, например, крайние величины скорости нормального поджелудочного отделения, то они часто относятся, как 1 : 20, а иногда это отношение делается еще гораздо большим. Подобный же результат дало и исследование различных порции одного и того же отделения на содержание бродил. С полным основанием этот ход отделения как всей массы пищеварительных соков, так и их бродил следует отнести на счет правильного изменения степени раздражения различных отделительных нервов в течение нормального пишеварения. А это изменение вполне понятно. Сначала, при поступлении пищи в пищевой канал и постепенном продвигании ее в более глубокие отделы канала, вследствие захватывания все большей и большей чувствительной поверхности, отраженное раздражение отделительных нервов все увеличивается, а затем начинает уменьшаться, когда при переваривании пищи ее первоначальный грубый вид сглаживается, а количество уменьшается вследствие всасывания. Итак, в начале пищеварения отделительные нервы раздражаются очень сильно, в дальнейших же ступенях все слабее и слабее. Изменения железистых клеточек оказываются в прямой связи с этим обстоятельством. Пока раздражение сильно, клеточки постепенно уменьшаются в объеме, т. е. разрушаются скорее, чем восстанавливаются. Когда же раздражение делается слабее, клеточки начинают расти, чтобы при минимальном раздражении достигнуть уже первоначального размера. Нужно заметить, что раздражение отделительных нервов, например поджелудочной железы, в нормальных условиях никогда не прекращается совершенно. После вышеизложенного понятно, что я в моих опытах имел такое преобладание разрушения над восстановлением. В этих опытах постоянно употреблялось только сильное раздражение; раз нерв вследствие утомления начинал действовать слабее, ток усиливался. Нужно ожидать, следовательно, что при искусственном слабом раздражении получится другой, больший процент восстановления. И я могу из моих опытов привести несколько данных в пользу такого заключения Как уже замечено выше, во втором ряде опытов больший процент восстановления сравнительно с первым рядом мог произойти именно потому, что в нем к периоду сильного искусственного раздражения присоединился период автоматического, более слабога и постепенно ослабевавшего до нуля раздражения. Еще более резко выступило это в тех двух опытах последнего ряда, в которых автоматическое раздражение продолжалось вплоть до конца опыта: 7--8 часов по прекращении искусственного раздражения. Выделилась масса слюны (до 150 г), а веса парных желез относились совершенно необычно: в одной паре работавшая железа весила лишь немногим меньше, чем неработавшая, а в другой паре - даже наоборот: работавшая была тяжелее покойной. Очевидно, хоть это и одиночные опыты, что при автоматическом раздражении восстановление даже преобладало над разрушением. Подобное же обстоятельство бросилось в глаза и при некоторых пробных опытах с раздражением язычных нервов. В тех из них, где эти нервы начинали, наконец, гнать слюну значительно даже и при слабых токах, оказывалось очень маленькое различие в весе парных желез. Конечно, толкование приведенных данных есть только вероятное, но не безусловно убедительное: мыслимо, что при автоматическом раздражении имеет определяющее значение не сила раздражения, а что-нибудь другое.
Заключение об особых отношениях разрушительных и восстановительных процессов в железе при сильных и слабых раздражениях находит себе еще поддержку и даже известное объяснение в особых свойствах нервов. Гейденгайном установлено, что при переходе слабого раздражения chorda tympani на сильное выделяется не только более слюны, но и более насыщенной. На основании этого факта, имея в виду гейденгайновское деление железыстых нервов на отделительные и трофические, нужно принять, что сильные токи относительно более раздражают трофические нервы, а слабые, наоборот, - отделительные. Если допустить, что отделительные нервы, обусловливая ток жидкости через железу, вместе с тем ведут и к отложению в железе нового вещества, a трофические нервы суть только разрушители железистой ткани, то мое заключение об отношении между силою раздражения и процентом восстановления уясняется даже в своем внутреннем основании. Последнее же допущение представляется очень правдоподобным. Существующие доводы за особые восстановительные, так называемые анаболические, нервы (Ленгли и Брэдфорд) были признаны мною недоказательными. Таким образом мое предположение об отделительных нервая как восстановителях и трофических как разрушителях железы, не имея ничего против себя, удобно бы связывало, даже объясняло как ход процессов в железах, наблюдаемых при нормальном пищеварении, так и результат сообщенных мною опытов.
Я кончил и теоретические рассуждения. Как сказано, они должны были иметь значение программы для дальнейших опытов. По этой программе опыты определения процента восстановления при разных силах раздражающего тока стоят на первом плане.
Я убежден, что продолжение и развитие опытов, подобных описанным, дадут твердую почву, на которой многообъемлющее и глубоко интересное учение о восстановлении живой ткани может разрабатываться плодотворно и далеко.
Иннервация желудочных желез у собаки
(С о в м е с т н о с Е. О. Ш м о в о й - С и м а н о вской)
Влияние нервной системы на деятельность желудочных желез. искони составляло предмет резкого разноречия между медициной и физиологией. Между тем как первая давно уже говорила о расстройствах отделительной иннервации желудка, вторая -- в лице лучших ее представителей - решительно отрицала всякую такую иннервацию. За последнее время в массе медицинских статей и целых книгах описываются отделительные неврозы желудка, причем существование иннервации желудочных желез разумеется само собою и высказывается только сетование, что физиология так отстала в этом отношении. И вместе с этих Гейденгайн первый современный авторитет в физиологии отделительной деятельности - свое изложение об иннервации желез желудка в «Руководстве к физиологии» Германа заканчивает следующими словами: «Результат многочисленных опытов гласит несомненно, что приходящие извне к желудку нервы не обладают никаким заметным влиянием прямого рода на отделение". Даже о возбуждающем влиянии местного механического раздражения слизистой оболочки желудка он находит возможным сделать два предположения: может быть, оно осуществляется при посредстве местного отделительного центра, заложенного в стенке желудка, но мыслимо также и непосредственное действие на отделительные элементы, в том роде, как это, по Дарвину, происходит у насекомоядных растений.
Наши опыты решают давний спор пользу медицины. Мы надеемся, что эти опыты заставят физиологов признать для желудочных желез те же иннервационные отношения, какие доказаны в настоящее время для многих других желез: слюнных, поджелудочной, потовых и т. д. Таким образом медицина вышла победительницею, и, конечно, не случайно. Едва ли можно не признавать, что вековечные и всегда глубока интересующие человека наблюдения болезненных явлений его организма должны были быть всегда ценным указанием относительно отправлений и их условий в различных частях тела. из всего клинического наблюдения поражения нервной системы в особенности должны были вести часто к плодотворным физиологическим заключениям, так как они представляют собой сплошь и рядом такие физиологические опыты, которые при современной физиологической технике даже и невозможно воспроизвести в лабораториях. И надо жалеть, что обмен между медициной и физиклогиею часто не так оживлен, как бы тому следовало быть [2].