Книги
Эмоциональный мозг
Подражательное поведение как пример компенсаторной функции эмоций на популяционном уровне
Переход к имитационному поведению чрезвычайно характерен для эмоционально возбужденного мозга. В сущности это частный случай доминантного реагирования на сигналы с малой (проблематичной) вероятностью их подкрепления, в данном случае — на сигналы, исходящие от других особей. Когда субъект не располагает данными или временем для самостоятельного и вполне обоснованного решения, ему остается положиться на пример других членов сообщества. Поскольку целесообразность приспособительных реакций относительна, имитационное поведение далеко не всегда является оптимальным. Показано, что мотивация следования за лидером при выборе дверцы в лабиринте сильнее у крыс, чем их собственный опыт. Крысы без лидера выбирают правильную дверь в 66 % случаев. Крысы, следующие за лидером, обученным выбирать противоположную дверь, делают правильный выбор только в 40 % [Konopasky, Telegdy, 1977]. В условиях массовой паники подражательное поведение может обернуться подлинной катастрофой. И все же в процессе длительной эволюции такое поведение, по-видимому, оказалось статистически выгодным и было закреплено естественным отбором.
Подкрепляющая функция эмоций на популяционном уровне: феномен эмоционального резонанса
Многократно показано, что животные способны реагировать на внешние проявления эмоционального состояния другой особи своего, а подчас — и другого вида. Более того, эти сигналы могут играть роль подкрепления при выработке инструментальных условных рефлексов, в том числе у крыс [Rice, Gainer, 1962; Greene, 1969].
Реакцию избегания при болевом раздражении другой особи мы вырабатывали у 267 взрослых белых крыс-самцов. В этих опытах было использовано экологически характерное для крыс предпочтение находиться в ограниченном пространстве. В установке, сконструированной инженером В. А. Пучковым (рис. 7), пол плексигласового «домика» представлял педаль, автоматически включавшую счетчик времени и болевое раздражение электрическим током лап другой крысы, находившейся за тонкой прозрачной звукопроницаемой перегородкой. Исследуемое животное один раз в день помещали в открытую, относительно просторную часть установки на 5 мин и регистрировали время его пребывания на педали. В течение 10 (в некоторых сериях опытов 5) дней вход в «домик» не сопровождался болевым раздражением второй крысы, в то время как на протяжении следующих 10 дней каждое появление исследуемого животного в «домике» приводило к включению тока силой 1–2 мА. Раздражение «жертвы» продолжалось 3–5 с, с пятисекундными интервалами, до тех пор пока исследуемая крыса находилась на педали.
Специально поставленные опыты со звуковой и зрительной изоляцией крыс показали, что в комплексе сигналов оборонительного возбуждения «жертвы» решающее значение для исследуемого животного имеет крик. Вот почему мы меняли партнеров в случайном порядке, а крыс — «жертв», склонных к пассивно-оборонительной реакции «замерзания», с хриплой или беззвучной вокализацией выбраковывали и заменяли другими. При угашении выработанной условной реакции избегания ток выключали, однако крыса — «жертва» по-прежнему оставалась в камере.
Оказалось, что на протяжении 2–3 (редко 6–8) опытов время пребывания на педали у 85 (то есть примерно у 32 %) исследованных животных становилось короче — 1 мин 30 с, а среднее число появлений в «домике» не превышало 3,1 (см. крысу № 13 на рис. 8). Некоторые крысы этой группы вообще перестали заходить в «домик», хотя большую часть времени проводили около входа на педаль. Среди остальных 182 крыс можно было выделить животных, находившихся на педали почти все 5 мин экспозиции, а также крыс, непрерывно перебегавших из открытого пространства в «домик» и выбегавших обратно при крике раздражаемой током «жертвы».
Нас интересовали три вопроса:
1) какие характерные черты присущи другим формам зоосоциального поведения крыс, различающихся по их реакциям на болевое раздражение партнера?
2) каким образом и в каких пределах можно изменить реакцию крыс на крик боли, издаваемый другой крысой?
3) повреждение каких отделов головного мозга и в каком направлении влияет на изучавшуюся нами реакцию избегания?
Ниже приводятся результаты опытов, призванных ответить на эти вопросы.
Ранее [Симонов, 1976] мы показали, что крысы, выработавшие реакцию избегания крика боли без предварительного знакомства с болевым раздражением, характеризуются высоким уровнем исследовательской активности в «открытом поле», отсутствием дефекаций и уринаций в ситуации «открытого поля» (показателя пассивно-оборонительной реакции страха), а также низкой агрессивностью при раздражении лап двух крыс электрическим током. Крысы, не обнаружившие способности к выработке условной реакции избегания, напротив, оказались малоактивными в «открытом поле», трусливыми (если судить по дефекациям и уринациям) и агрессивными. Сочетание этих трех качеств наиболее неблагоприятно для выработки условного рефлекса.
В опытах, поставленных Д. З. Партев[1], у каждой из 10 крыс отдельно вырабатывали условный оборонительный рефлекс на звонок, подкрепляемый болевым раздражением лап. Камера с решетчатым полом была разделена на две половины перегородкой, имевшей дверцу Электрический ток подавали на решетку пола одной половины камеры после 5–8 с изолированного действия звонка, который продолжался еще 10 с вместе с болевым раздражением.
После того как у всех 10 крыс был выработан стабильный оборонительный условный рефлекс (80-100 % реакций на условный сигнал), в камеру помещали одновременно от 3 до 5 животных. Если теперь включить звонок, то крысы, ранее быстро перебегавшие на безопасную половину камеры, будут вести себя по-разному. Одни из них по-прежнему устремляются на безопасную половину, в то время как другие начинают драться. Некоторые из крыс, перебежав на безопасную половину, встают в дверях и атакуют крыс, пытающихся спастись от тока. Подчеркнем, что убегают или дерутся одни и те же животные. Даже в том случае, если условный сигнал (звонок) застает агрессивную крысу рядом с открытой дверцей, крыса не убегает от тока, а направляется в прямо противоположном направлении, чтобы атаковать другое животное. При выработке условной реакции избегания крика боли (до начала экспериментов с групповым поведением) у крыс, находившихся в «домике» в среднем 2,13 мин, в групповом эксперименте после 120 применений условного сигнала (звонка) в 80 % случаев зарегистрирован уход на безопасную половину и только в 14 % —драки. У тех же крыс, которые находились в «домике» в среднем 4,03 мин, количество драк достигло 65 %, а уход от раздражения током наблюдался лишь в 46 % случаев. Что касается «защиты безопасной территории», то эта форма поведения наблюдалась у обеих категорий крыс в одинаковом числе случаев (6 %). Опыты Д. P. Партев показали, что характер реакции на болевое раздражение партнера представляет устойчивую черту индивидуальных особенностей данного животного, закономерно коррелирующую с его поведением в ситуации угрозы собственного болевого раздражения, причем крысы малочувствительные к крику боли «жертвы» более склонны к агрессивному поведению.
Можно ли изменить реакцию крыс на крик боли другой особи и в каких пределах?
182 крысы, находившиеся на педали свыше 1 мин 30 с, мы от 3 до 10 раз использовали в качестве «жертв» при выработке условной реакции избегания у их партнеров. Эта процедура привела к тому, что у 14 животных время пребывания в «домике» резко сократилось, хотя у 68 оно осталось без изменений (см. крыс № 11,12 на рис. 8). Можно было предположить, что применение тока ведет к выработке обычного оборонительного рефлекса, где ограниченное пространство становится условным сигналом болевого раздражения. Экспериментальные факты противоречат подобному объяснению. Как только начинается угашение условного рефлекса (то есть крысу-«жертву» перестают раздражать током), крысы уже в первой пробе находятся на педали свыше 4 мин. Значит, использование подопытного животного в качестве «жертвы» ведет не к выработке обычного оборонительного условного рефлекса, а повышает чувствительность к сигналам оборонительного возбуждения другой особи. Аналогичные результаты ранее получил Р. Чёрч [Church, 1959].
Впрочем, процедура воздействия электрическим током дает этот эффект только у 42 % от общего количества исследованных животных. 68 из 267 крыс (или 26 %) продолжали находиться на педали свыше 3 мин (в среднем 3,77 мин) и после ознакомления с током. Возник вопрос: не объясняется ли эта «нечувствительность» к крику боли другой особи затруднением в выработке условной реакции избегания, то есть дефектом механизмов научения, приобретения новых навыков? Известные основания к такому предположению у нас возникли во время опытов Д. 3. Партев, где выяснилось, что крысы, находившиеся на педали меньше 2 мин, реагируют на сигнал раздражения током условным оборонительным рефлексом в 81 % всех применений этого сигнала на протяжении 11–14 дней, а крысы, находившиеся на педали свыше 3 мин, — только в 61 % случаев. Вот почему нас заинтересовало предложение д-ра В. Вецель[2] применить оротовую кислоту — вещество, способствующее консолидации вновь выработанных условных связей.
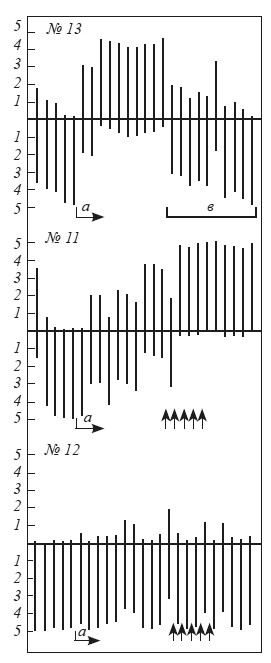
Рис. 8. Динамика выработки реакций избегания у крыс № 11, 12, 13
Абсцисса — последовательные пробы. Ниже оси ординат — время пребывания крысы в «домике» (мин), выше — время пребывания вне «домика»;