Книги
Неандертальцы: какими они были, и почему их не стало
Следовательно, будем ли мы относить неандертальцев и людей современного анатомического типа к разным видам, или включать их в качестве подвидов в один вид, это не может иметь никаких последствий для решения вопроса о возможности гибридизации между ними. Я думаю, что даже если бы мы точно знали, что потенциально эти две формы были способны к скрещиванию и между ними происходил обмен генами, то и тогда их следовало бы рассматривать и обозначать как отдельные виды. Ведь по многим признакам, включая качественные и метрические характеристики черепа, они различаются между собой больше, чем современные люди разных рас, а также гориллы, шимпанзе, павианы и макаки разных подвидов и видов.
«Неандертальцы весьма своеобразны по строению их скелета и, особенно, по форме черепа. Если вы посмотрите на скелеты любых двух близкородственных видов ныне живущих приматов (например, черного и коричневого лемуров), вы обязательно обнаружите, что различия между ними намного меньше тех, что отделяют скелет типичного неандертальца от нашего собственного. Если исходить из установленных стандартов систематики млекопитающих, то ясно, что неандертальцы — это отдельный вид...», — уверен американский антрополог И. Таттерсол (Tattersall 1995: 10). Эту уверенность разделяют с ним многие его коллеги.
«Если бы неандертальцы и современные люди были грызунами или антилопами, любой специалист по палеонтологии позвоночных, не колеблясь, отнес бы их к разным палеонтологическим видам», — пишет французский исследователь Ж.-Ж. Ублэн. В реальности, однако, продолжает он, «речь идет о людях, причем людях с очень схожими формами технических и поведенческих адаптаций. В прошлом именно это было одной из причин, побуждавших антропологов объединять тех и других в один вид» (Hublin 2006: 57).
Кроме анатомических различий и, по меньшей мере, полумиллиона лет самостоятельной эволюционной истории, в пользу обособленного положения неандертальцев по отношению к гомо сапиенс свидетельствуют и генетические данные. Выше мне уже приходилось несколько раз ссылаться на сведения, добытые благодаря появившейся в последние годы чудесной возможности заглянуть в ДНК давно вымерших видов. Теперь пришло время рассказать об этом подробней.
Палеогенетике от роду — четверть века. Первая успешная попытка извлечь, реконструировать и проанализировать ДНК из мертвых тканей была осуществлена в США в 1984 г., причем «тканям» этим — шкуре вымершей к тому времени разновидности зебры под названием квагга — было всего-то 140 лет. Затем пришел черед египетских мумий и палеоиндейских скелетов возрастом в несколько тысяч лет, а сегодня вполне обычным делом стало уже изучение генов людей и животных, живших десятки тысяч лет назад. Чуть ли не каждую неделю в специальных журналах появляются статьи с новой информацией о ДНК мамонтов, пещерных медведей и прочих экзотических существ. С 1997 г. в число этих прочих входят и неандертальцы.
Прорыв, начало которому было положено двадцать пять лет назад, стал возможен благодаря методу, именуемому полимеразной цепной реакцией. Этот метод, изобретенный американским биохимиком К. Муллисом в 1983 г., позволяет получить неограниченное количество пригодных для анализа копий фрагментов ДНК — как современной, так и древней. Сопоставляя последовательность нуклеотидов во фрагментах ДНК, добытых из костей (или, скажем, слюны, если речь идет о доселе здравствующих представителях животного мира) разных индивидов и/или видов, можно оценить генетическое расстояние между ними, определить, кто из сравниваемых друг другу близкая родня, а кто — седьмая вода на киселе, и даже рассчитать — пусть и очень приблизительно — время, когда жил их последний общий предок.
Все перечисленное проделали и с ДНК неандертальцев. Результаты получились очень интересные. Однако прежде чем познакомиться с ними, нам следует, наверно, вспомнить некоторые азы биологии. Ну, хотя бы для того, чтобы у читателей, успевших несколько подзабыть школьный курс этой науки, не появилось подозрение, что их просто пытаются водить за нос с помощью непонятных слов.
Итак, ДНК. Эту аббревиатуру генетики придумали, чтобы не мучиться по сто раз на дню, выговаривая или набирая на клавиатуре компьютера слова «дезоксирибонуклеиновая кислота». Молекулы, или, точнее, макромолекулы ДНК — место хранения генетической информации, определяющей индивидуальные особенности и характер развития каждого организма и передаваемой от поколения к поколению. Каждая такая макромолекула — это цепочка, образованная двумя тяжами (нитями), спирально закрученными один относительно другого, а каждый тяж представляет собой последовательность тысяч и миллионов нуклеотидов. Эти нуклеотиды были бы похожи между собой, как близнецы, если бы не так называемые азотистые основания, входящие в их состав наряду с молекулой сахара (дезоксирибоза) и фосфата (фосфорная кислота), и представленные четырьмя разными типами. Два типа — гуанин и аденин — называются пуриновыми основаниями, а еще два — тимин и цитозин — пиримидиновыми.
Молекулы сахара и фосфата образуют остов тяжей (нитей) ДНК, а основания находятся между тяжами и посредством слабых водородных связей соединяют между собой противолежащие нуклеотиды (рис. 18). При этом аденин может соединяться только с тимином, а гуанин только с цитозином. Последовательность этих парных оснований — шифр, в котором закодированы свойства белковых молекул и, в конечном счете, свойства всего живого. Гены, т. е. сегменты ДНК, отвечающие за синтез разных белков, могут иметь длину от нескольких десятков до нескольких миллионов парных оснований.
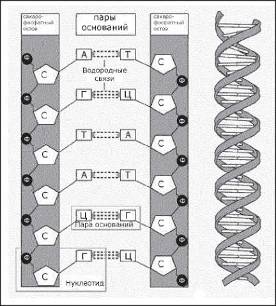
Рис. 18. Структура двойной спирали ДНК. Буквами обозначены: Ф — фосфат, С — сахар, А — аденин, Т — Тимин, Г — гуанин, Ц — цитозин.
Иногда вследствие слабости водородных связей при репликации ДНК происходят мутации, т. е. «перестановки» оснований, или, иными словами, изменения в порядке последовательности нуклеотидов. В большинстве своем такие мутации нейтральны по отношению к естественному отбору. Они не выбраковываются и не подхватываются им, поскольку не отражаются на приспособленности организмов, и благодаря этому обстоятельству скорость их накопления на молекулярном уровне в целом постоянна. Следовательно, если для истории изучаемой группы организмов имеются более или менее четкие и надежно датированные палеонтологические реперы (точки отсчета), то эту скорость можно рассчитать. Например, для гоминид, а иногда и для всех обезьян вообще в качестве основы расчетов используется генетическое расстояние (т. е. количество различий в последовательности нуклеотидов в ДНК) между современными людьми и шимпанзе, эволюционные пути которых, судя по ископаемым находкам, разошлись около 6 млн. лет назад.
Зная скорость накопления мутаций и генетическое расстояние между разными особями или таксонами (т. е. видами, родами и т. д.), можно не только судить о близости их родства, но и рассчитывать время дивергенции, расхождения от общего предка, применяя для этого упоминавшийся уже выше метод молекулярных часов. Обычно для таких расчетов используют ДНК из митохондрий клеток, которая, в отличие от ДНК, содержащейся в ядрах, представлена в каждой клетке сотнями и тысячами идентичных копий, наследуется только по женской линии, и к тому же характеризуется более высокими темпами накопления мутаций.
На основании изучения митохондриальной ДНК были получены и первые сведения о генетике неандертальцев. Сначала, в 1997 г., удалось выделить пригодный для анализа фрагмент мтДНК из одной из костей, найденных в 1856 г. в гроте Фельдгофер, и сравнить его с аналогичным участком ДНК современного человека. Следующими «поставщиками» ископаемого генетического материала стали кости из пещер Виндия (Хорватия) и Мезмайская (Северный Кавказ, Россия). К настоящему времени генетические данные имеются уже примерно по 15 неандертальцам, жившим или погребенным в разное время в пещерах Испании (Эль Сидрон), Франции (Ля Шапелль-о-Сен, Рош де Вилленев), Бельгии (Анжи, Складина), Италии (Монте Лессини), Узбекистана (Тешик-Таш) и Южной Сибири (пещера Окладникова).
В последние годы палеогенетики все больше внимания стали уделять изучению ядерной ДНК. Это и не удивительно, поскольку именно в ней сосредоточена львиная доля генетической информации.[8] В 2006 г. группа исследователей из Института эволюционной антропологии им. Макса Планка в Лейпциге объявила даже о предстоящей в скором времени полной расшифровке неандертальского ядерного генома, и хотя сейчас уже очевидно, что названный тогда срок выполнения проекта — два года — был чересчур оптимистичным, работа идет, и идет, судя по поступающим сообщениям, вполне успешно. По оценке одного из ее инициаторов и руководителей, С. Пээбо, к началу 2009 г. было «раскодировано» около 60 % ядерной ДНК неандертальцев (а точнее, двух неандерталок) из Виндии, а когда моя книга дойдет (надеюсь) до читателя, этот показатель, возможно, будет уже приближаться к 100 %.
Что же касается митохондриального генома, то здесь стопроцентный результат был достигнут еще в 2008 г. Исходным материалом тоже послужила кость из Виндии, имеющая прямую (т. е. полученную по самой этой кости, а не по сопровождающим ее находкам) радиоуглеродную дату 38,3+2,1 тыс. лет назад. Исследование мтДНК, выделенной из этой кости, позволило идентифицировать и расставить по местам все 16565 пар нуклеотидных оснований, составлявших кольцевую молекулу митохондриальной дезоксирибонуклеиновой кислоты неандертальца[9], и сравнить полученную таким образом последовательность с последовательностями, выявленными у 53 современных людей разной расовой принадлежности. При попарном сопоставлении оказалось, что из шестнадцати с половиной тысяч позиций у нас и неандертальцев не совпадают в среднем 206, тогда как геномы современных людей различаются между собой гораздо меньше: максимум по 118 позициям, минимум по 2 (Green et al. 2008). Эти данные в целом подтверждают выводы первых палеогенетических работ, согласно которым разница в последовательности нуклеотидов в мтДНК неандертальцев и современных людей примерно в три или три с лишним раза превышает «генетическое расстояние» между ныне существующими расовыми группами человека. Много это или мало? Это достаточно, чтобы продолжать рассматривать неандертальцев как особый вид, но совсем недостаточно, чтобы вообще не считать их за людей. Ведь если провести аналогичное сравнение, например, с мтДНК шимпанзе, то выяснится, что от них мы отличаемся в среднем по 1500 позициям.
Генетический материал был получен также из костей гомо сапиенс ранней поры и середины верхнего палеолита. Сравнение одних и тех же участков цепей ДНК поздних неандертальцев, верхнепалеолитических гомо сапиенс и современных людей показало, что, если между последними двумя группами, несмотря на хронологический разрыв в 20 с лишним тысяч лет, нет сколько-нибудь заметных различий, то неандертальцы явно стоят особняком. В частности, от людей из грота Пальиччи, живших 23–25 тыс. лет назад, их отделяет такая же генетическая дистанция, как и от современных людей. Ни один из изученных до сих пор образцов ДНК гомо сапиенс — современных и палеолитических — не дал последовательности, типичной для неандертальцев, а все неандертальские образцы, из какой бы части Европы они ни происходили, оказались в целом довольно близки между собой (Lalueza-Fox et al. 2006; Orlando et al. 2006; Briggs et al. 2009).
Таким образом, палеогенетические данные, как и анатомические, говорят в пользу обособленного видового статуса неандертальцев. Большинство оценок времени происхождения этого вида (или, точнее говоря, времени, когда жил последний общий предок всех «классических» неандертальцев), полученных методом молекулярных часов, укладываются в интервал от 150 до 250 тыс. лет назад, а с учетом стандартного отклонения от 120 до 350 тыс. лет назад. Разделение линий неандертальцев и современных людей произошло, судя по первым результатам применения того же метода, скорее всего, где-то в интервале от 500 до 700 тыс. лет назад, или, если брать крайние даты, от 320 до 850 тыс. лет назад (Green et al. 2006; Noonan et al. 2006). Наконец, самые последние расчеты времени этого события, основанные на данных по полностью расшифрованной мтДНК из Виндии, дали дату 660+140 тыс. лет (Green et al. 2008). Она хорошо согласуется с палеоантропологическими данными и является, вероятно, наиболее реалистичной.
Глава 5. Дети севера
Почему неандертальцы стали такими, какими они стали? Что привело к появлению у них тех анатомических особенностей, которые отличают их от гомо сапиенс и других гоминид и позволяют считать их особым биологическим видом? Вопрос этот очень труден, но не неразрешим. Антропологам есть что на него ответить. Ответ, конечно, будет далеко не исчерпывающим, но, как ни странно, гораздо более полным, чем в том случае, если бы мы поставили тот же вопрос применительно к своему собственному виду. Хотя очень многие специфические черты неандертальской морфологии пока еще не получили сколько-нибудь удовлетворительного объяснения, причины и общий функциональный смысл эволюции неандертальцев понятны сегодня все же лучше, чем причины и смысл большинства тех анатомических изменений, которые сопутствовали появлению гомо сапиенс.
В целом, конечно, мало кто сомневается в том, что направление биологической эволюции обоих видов, особенности их анатомии и образа жизни в значительной мере зависели от природных условий, в которых им приходилось жить и к которым они должны были приспосабливаться. Однако если характер задач, которые окружающая среда могла ставить перед неандертальцами, более или менее ясен, то о гомо сапиенс этого пока не скажешь. Ведь становление первых происходило на севере, в краю с довольно суровой, переменчивой и очень требовательной природой, а вторые формировались в тропиках, где климат неизмеримо мягче, а перепады его далеко не столь резки и непредсказуемы, как в высоких широтах. К разговору о гомо сапиенс нам еще предстоит вернуться в одной из следующих глав, а сейчас попытаемся представить, каковы были естественные условия существования неандертальцев, и в чем конкретно сказалось влияние этих условий на их анатомию и внешний облик?