Книги
Эволюция человека. Книга II. Обезьяны, нейроны и душа
Обобщив все доступные данные, на основании которых можно судить о динамике увеличения ОКРП с возрастом у человека и других обезьян, Рид пришел к выводу, что разнообразные когнитивные способности, предположительно отражающие величину ОКРП, раньше всего начинают развиваться у людей, позже всего – у низших (нечеловекообразных) обезьян; человекообразные обезьяны занимают промежуточное положение. Скорость, с которой развиваются эти способности, максимальна у человека, минимальна у низших обезьян. Наконец, завершение развития этих способностей происходит раньше всего у низших обезьян, позже всего – у людей; человекообразные, как всегда, посередине.
Таким образом, у людей умственное развитие начинается раньше, идет быстрее и заканчивается позже, чем у других обезьян. В целом интеллектуальное развитие человека и шимпанзе остается более или менее сравнимым примерно до трехлетнего возраста. После этого развитие шимпанзе резко затормаживается, и люди начинают их стремительно опережать. Для шимпанзе все заканчивается в возрасте около четырех лет при уровне ОКРП = 2 или, самое большее, 3. Люди же продолжают развиваться по прежней «траектории», достигая уровня ОКРП ≈ 7 примерно к 12 годам.
Крайне интересно было бы узнать, хотя и не совсем понятно, какие эксперименты нужно поставить для получения ответа на подобный вопрос, как бы работало человеческое мышление и как была бы устроена наша речь, в особенности ее грамматическая структура (здесь, к слову, стоит упомянуть и о том, что, согласно известной теории Ноама Хомского, в честь которого не без иронии был назван вышеупомянутый шимпанзе Ним Чимпски, у человека имеется «врожденная грамматика» – некое генетически обусловленное представление о грамматической структуре речи, хотя эта теория в настоящее время не разделяется большинством экспертов), и как развивалась бы человеческая культура и наука, если бы наша эволюция не остановилась на достигнутом уровне развития кратковременной рабочей памяти, соответствующем, по мнению большинства психологов, ОКРП ≈ 7, хотя некоторые, как отмечалось выше, предпочитают более осторожно говорить о величине 7 ± 2, о чем можно прочесть в недавно вышедшей книге известного психолога и нейробиолога Криса Фрита «Мозг и душа»
Развивая свои идеи, Рид проанализировал развитие палеолитических технологий, а также увеличение размеров мозга, и попытался по этим косвенным признакам выяснить, как менялся в ходе антропогенеза объем кратковременной памяти. Технологии изготовления орудий Рид делит на семь групп по уровню «концептуальной сложности»: от использования готовых палок, от которых нужно только оторвать лишние сучки и листья (уровень 1), до верхнепалеолитической технологии последовательного отщепления множества одинаковых лезвий от одного и того же призматического ядра (уровень 7). По мнению Рида, только технологии уровня 7, появившиеся менее 50 тыс. лет назад, бесспорно являются рекурсивными. Их рекурсивность состоит в том, что лезвия отщепляются не как попало, а с таким расчетом, чтобы одновременно подготовить ядро для отщепления следующего лезвия. При этом нужно одновременно держать в голове трехмерную форму ядра, контролировать его позицию и с большой точностью манипулировать отбойником. Технология шестого уровня – леваллуазское расщепление, появившееся свыше 700 тыс. лет назад, но широко распространившееся много позже, в среднем палеолите, – возможно, тоже требовала рекурсивного мышления, но в этом Рид не совсем уверен. Другие авторы указывают на необходимость высокоразвитой ДРП – но не КРП – для овладения леваллуазским мастерством (см. главу «От эректусов к сапиенсам» в кн. 1).
Рид предполагает, что у
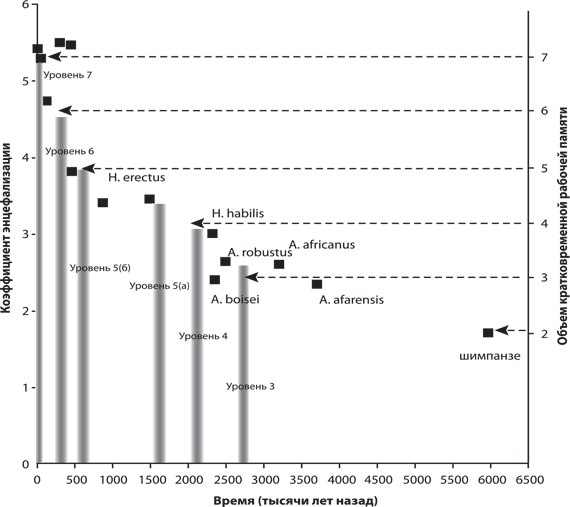
Гипотеза Рида выглядит довольно правдоподобно, хотя в ней есть ряд слабо проработанных моментов (например, не очень четко аргументирована связь между величиной ОКРП и способностью к рекурсивному мышлению). Однако генеральная идея о том, что уникальность человеческого интеллекта во многом определяется увеличенным объемом памяти, в том числе кратковременной рабочей, скорее всего верна.
Помимо прочего, большой ОКРП должен повышать новаторско-изобретательский потенциал
Глава 2
Душевная механика
Универсальный аппарат для принятия решений
Психика, она же душа, является результатом работы мозга. Мозг сделан из нервных клеток – нейронов. Мы не будем вдаваться в тонкие детали устройства нервных клеток, ведь эта книга не учебник по нейробиологии. Но несколько базовых фактов все-таки придется привести, потому что без них трудно понять нашу душевную механику.
Нейрон – универсальное живое устройство для принятия решений. Это главное, что нам следует о нем знать.
Я чуть было не назвал его простейшим или элементарным устройством, но вовремя вспомнил, что есть и более простые биологические структуры, способные к принятию решений: разнообразные биохимические и генетические «переключатели»
Два слова о строении нейрона. У него есть центральная толстенькая часть – «тело», в котором находится клеточное ядро с генами. От тела отходят два вида отростков: «входные» (дендриты) и «выходные» (аксоны). Дендритов обычно много, а аксон, как правило, один, но на конце он может ветвиться.

Главная задача дендритов – сбор информации. Они могут получать сигналы от специальных белков-рецепторов (например, обонятельных, вкусовых или светочувствительных), реагирующих на факторы внешней или внутренней среды, и в этом случае нейрон называется сенсорным. Но в большинстве случаев дендриты получают сигналы от других нейронов, чаще всего – от их аксонов. Для того чтобы обмениваться сигналами, нейроны используют специальные вещества –
Как правило, передача сигнала от аксона одного нейрона к дендриту или иной части другого нейрона осуществляется в специальной контактной зоне, которая называется синапсом. Главные составные части синапса – это
Кроме обычных, химических синапсов, в которых сигнал от нейрона к нейрону передается при помощи нейромедиаторов, бывают еще электрические синапсы, но они играют менее важную роль, и в нашем рассказе мы постараемся без них обойтись.
